The Species Composition of Phytoplankton and Nutrient Content in the Nipa Nypa fruticans Ecosystem on the West Coast of Aceh, Indonesia

Dewi Fithria | Hairul Basri | Indra Indra | Zainal A. Muchlisin*
© 2022 IIETA. This article is published by IIETA and is licensed under the CC BY 4.0 license (http://creativecommons.org/licenses/by/4.0/).
OPEN ACCESS
Nipa Nypa fruticans grows in estuaries that are affected by sea waters and freshwater. Therefore, the Nipa habitat develops into an area of water mass circulation and becomes rich in nutrients that trigger the growth of phytoplankton as the primary producer in the aquatic ecosystem. This study aimed to examine the species composition and distribution of phytoplankton and nutrient content in the nipa ecosystem on the west coast of Aceh, Indonesia. It was conducted from July 2018 to September 2019 and sampling was carried out in two areas, namely Kuala Bubon Aceh Barat district and Kuala Tadu, Nagan Raya district. A total of 100L of surface water samples was taken from three points at each location. Subsequently, the water was filtered with a plankton net mesh No. 25 and preserved with a 4% Lugol solution. The water sample from both locations was also analyzed for nitrogen and phosphorus contents. The results showed that there were 20 species of phytoplankton in both sampling locations, with Kuala Bubon consisting of 13 species with density of 160.87 cells L-1, and 15 species in Kuala Tadu with density of 273.64 cells L-1. Haramonas sp. was the dominant species in Kuala Bubon, while Amphisolenia bidentata was dominant in Kuala Tadu. Based on the nutrient content (nitrate+nitrite and phosphate), the average nutrient content in Kuala Bubon was 1.31 mg L-1 (mesotrophic) and 0.12 mg L-1 phosphate (hypertrophic). Meanwhile, Kuala Tadu had 1.078 mg L-1 nitrate+nitrite (mesotrophic) and phosphate, 0.22 mg L-1 (hypertrophic) waters. These findings are an early indication that the estuary of Kuala Bubon and Kuala Tadu have been contaminated by organic matter.
Nypa fruticans, organic matter, primary producer, phyoplankton, water productivity
Aceh province has a great potential for aquatic biological resources, and the nipa ecosystem is one of these resources in this province, situated in the western region of Indonesia. nipa is a member of the palm group and typically grows alongside mangroves. Furthermore, sea water and fresh water from the mainland have an impact on the nipa ecosystem, making it an area of water mass circulation and high nutrients that are needed by plankton, particularly phytoplankton to grow and develop, resulting in high water productivity [1, 2].
Phytoplankton are microscopic organisms that inhabit various types of waters, such as freshwater, brackish, and seawater, and whose movements are dependent on currents [3]. Phytoplankton are autotrophs that act as primary producers in aquatic ecosystems [4]. Therefore, phytoplankton influence the survival and growth of aquatic organisms [3, 5].
Based on field observations along the west coast of Aceh, Indonesia, nipa grows in several areas such as Kuala Bubon, West Aceh and Kuala Tadu, Nagan Raya. Its habitat in Kuala Bubon and Kuala Tadu has an area of approximately 166 ha and 23 ha, respectively. Both of these ecosystems are categorized as semi-enclosed waters that tend to be shallow, narrow, and notched due to their geomorphological characteristics of bar-built estuaries [6, 7].
The enrichment of organic matters from upstream and downstream of the estuary results in high levels of nutrients, particularly nitrate and phosphate which can trigger eutrophication, thereby reducing the quality of the estuary waters where nipa live [8, 9]. This condition changes the composition, distribution, and abundance of phytoplankton and possibly the occurrence and blooming of toxic species in estuary waters. Nutrient variation in estuary waters is strongly related to the distribution and abundance of phytoplankton in estuaries, particularly the nipa ecosystem [2, 10], and these can be used as a bioindicator for waters productivity [11].
Currently, the estuary waters of Kuala Bubon and Kuala Tadu are threatened by ecological perturbation due to the conversion of the nipah ecosystem into the settlement, agriculture, and fisheries areas. The dominant activities in the two locations are fish landing, and fish market, where wastewater from these sources is directly discharged into the waters without being treated. It was assumed that these domestic activities along the two estuaries have the potential to pollute these waters. In this case, phytoplankton is the first aquatic organism to respond to these impacts.
Previous studies on the diversity and abundance of phytoplankton as a bioindicator in several Indonesian waters have been conducted. For instance, Muchlisin [12] studied the diversity and abundance of phytoplankton as a biological indicator of damage and pollution in the Sarah River, Aceh Besar, Aceh Province; Unbekna et al. [13] studied the plankton abundance and diversity in the Northern part of Lembata Island, Indonesia. Rahayu et al. [14] reported the plankton community structure and trophic status of Wadaslintang Reservoir in Indonesia. The abundance of phytoplankton in Sugihan River, South Sumatra was reported by Mulyadi et al. [15], and Jannah and Muchlisin [16] have been evaluated the phytoplankton community at estuary of Krueng Aceh River post tsunami. However, the previous studies were generally carried out in rivers, lakes, and estuaries without the nipa plant, while studies on the abundance and distribution of phytoplankton in the estuary associated with the nipa ecosystem, particularly on the West coast of Aceh have never been reported.
Information on the diversity and distribution of phytoplankton is crucial to document concerning to determine the productivity status of the waters and strategize a better management plan. Currently, the productivity status especially the phytoplankton diversity and nutrition content of the Kuala Bubon and Kuala Tadu estuaries were not available. Hence, the objective of this study was to assess the species composition and distribution of phytoplankton and nutrient content in the nipa ecosystem on the west coast of Aceh, Indonesia.
This is the first report on the species composition of phytoplankton in the nipa ecosystem from Indonesia. Therefore, this data is very important as baseline data for water monitoring programs in the future. The study was conducted in two location along the western coast of Aceh, Indonesia, namely; Kuala Bubon and Kuala Tadu. The data were collected from July to September 2019. The findings showed that the nitrate + nitrite content in both Kuala Bubon and Kuala Tadu were classified as mesotrophic waters, while based on phosphate content both locations were classified as hypertrophic water. However, the Kuala Bubon and Kuala Tadu had low species richness and low abundance of phytoplankton. The high content of nitrate and phosphate causes eutrophication which suppressed species richness as recorded in this study.
2.1 Description of the study site
This study was conducted from July 2018 to September 2019 in Kuala Bubon, West Aceh Regency and Kuala Tadu, Nagan Raya Regency; where the nipa plant grows. Purposive sampling was used to determine three sampling stations in each area; Station 1 was located near the river mouth; Station 2 was located in the middle of the estuary, and Station 3 was at the end of the estuary waters leading upstream. Subsequently, three sub-stations [sampling points] were determined randomly. The GPS coordinates of every sampling location are as follows: Kuala Bubon station: (St. I): 04°13'27.02", 096°03'33.87"; (St.II): 04°12'45.13", 096°03'05.73"; (St. II): 04°12'51.08", 096°02'46.20"; Kuala Tadu: (St. I): 04°01'11.16", 096°16'23.00"; (St. II): 04°01'27.73", 096°16'13.40"; (St. III): 04°01'44.09", 096°16'05.44", as shown in Figure 1.
Figure 1. The map of the study area showing the sampling locations (Red dots are in Kula Tadu, green dots are in Kuala Bubon)
2.2 Water sampling
A total of 100L of water was collected using a plastic bucket and filtered using a size 25 plankton net, then 50 ml of filtered water sample was put into the sample bottle and two drops of 4% Lugol's solution as a preservative were added and mixed homogeneously. The water samples were then transported to the Integrated Laboratory of Teuku Umar University for further analysis. The Identification of phytoplankton was conducted based on guideline books, including Davis [17], Prescott [18], and Yamaji [19]. The measurement of water quality parameters such as temperature, turbidity, salinity, pH, and dissolved oxygen was carried out in-situ, while chemical oxygen demand (COD), nitrate, nitrite, and phosphate were measured ex-situ.
2.3 Estimation of phytoplankton abundance
The abundance of phytoplankton was calculated by the sweep method as follow; one drop of the water sample was placed on a glass slide, covered with a cover glass, and then observed with a stereo microscope (SMT 4/SMC /Trinocular Tube 1.6) at a magnification of 100X. The phytoplankton abundance was calculated based on APHA [20] as follows:
$\mathrm{N}=Z\left[\frac{X}{Y}\right]\left[\frac{1}{V}\right]$
where, N= total abundance of phytoplankton (cells L-1), Z= total individual of respective species of phytoplankton, X= total volume of filtered water sample (50 ml), Y= water volume observed onto object glass (0.06 ml), V= Total volume of water sample (100 L).
2.4 Nitrate and phosphate analysis
The nitrate and nitrite contents in the waters were analyzed based on the standard procedure proposed by Sandu et al. [21], while the phosphorous content in the water was analyzed based on Eisenreich et al. [22].
2.5 Data analysis
Data on species composition, distribution, and water quality were presented in tables and figures and then analyzed descriptively by comparing the results with previous reports and other relevant references.
A total of 20 species of phytoplankton belonging to six classes were recorded in both locations, with five species in Kuala Bubon, seven species in Kuala Tadu, and eight species found in both locations, as shown in Table 1. Based on species richness, Dinophycea was predominant in Kuala Bubon, and Bacillariophyceae was predominant in Kuala Tadu. Meanwhile, based on the total individual, Haramonas sp. and Leptocylindrus danicus were the predominant species in Kuala Bubon, while Amphisolenia bidentate and Haramonas sp. were the predominant in Kuala Tadu. In addition, the density of phytoplankton was 160.87 cells L-1 in Kuala Bubon, and 273.64 cells L-1 in Kuala Tadu.
Table 1. Species composition and distribution of phytoplankton in the nipa N. fructicans ecosystem at Kuala Bubon and Kuala Tadu estuaries, Aceh Province, Indonesia
|
Class |
Species |
Total individual [cell L-1] |
|
|
KB |
KT |
||
|
Bacillariophyceae |
Aulacoseira sp. |
9.95 |
18.24 |
|
|
Delphineis surirella |
- |
3.32 |
|
|
Diatom |
9.95 |
33.17 |
|
|
Leptocylindrus danicus |
33.17 |
4.98 |
|
|
Leptocylindrus menimus |
6.63 |
1.66 |
|
|
Nitzschia rostrata |
- |
4.98 |
|
Dinophyceae |
Amphisolenia bidentata |
18.24 |
107.79 |
|
|
Ceratium fuscus |
- |
6.63 |
|
|
Ceratium hirundinella |
- |
6.63 |
|
|
Gonyaulax spinifera |
- |
1.66 |
|
|
Gymnodinium heterostratum |
- |
1.66 |
|
|
Gyrodinium instriatum |
1.66 |
- |
|
|
Dinophysis acuminata |
- |
1.66 |
|
|
Dinoflagelata sp. |
16.58 |
19.90 |
|
|
Dinophysis caudata |
4.98 |
6.63 |
|
Raphidophyceae |
Haramonas sp. |
53.07 |
51.41 |
|
Coscinodiscophyceae |
Dactyliosolen blavianus |
3.32 |
- |
|
|
Melosira sp. |
1.66 |
1.66 |
|
Cyanophyceae |
Trichodesmium erythraeum |
- |
1.66 |
|
Gonyaulacaceae |
Schuetiella mitra |
1.66 |
- |
|
Total cells |
|
160.87 |
273.64 |
|
Total species |
|
12 |
17 |
Note: KB= Kuala Bubon, KT= Kuala Tadu
Hence, Haramonas sp. is predominant in both locations, and the abundance of species was almost similar in these locations. Currently, there are only two species of Haramonas are known, namely H. dimorpha and H. pauciplastida [23]. However, of the 20 phytoplankton species recorded in this study, A. bidentata was the most abundant in Kuala Tadu. However, this species is also found in Kuala Bubon but the numbers do not dominate. According to Daugbjerg [24], A. bidentata is belonging to the Dinophyceae group, which is marine phytoplankton that widely distributed in tropical and subtropical regions. According to Gul and Saifullah [25], this species was also predominant in the water of the North-West of Arabian Sea Shelf of Pakistan.
Water quality measurements showed that in Kuala Bubon, temperatures ranged from 22℃ to 26℃, salinity ranged from 15 ppt to 20 ppt, water brightness ranged from 30 cm to 50 cm, dissolved oxygen ranged from 5.4 to 6.0 mg L-1, pH from 6.6 to 7.2, chemical oxygen demand ranging from 45.05 to 53.25 mg L-1. While, at Kuala Tadu, the temperature ranged from 23℃ to 27℃, salinity ranged from 14 ppt - 20 ppt, water brighness ranged from 30 to 40 cm, dissolved oxygen 5.6 to 6.0 mg L-1, pH ranged from 6.6-6.9, chemical oxygen demand ranged from 40.96 to 49.15 mg L-1, as shown in Table 2. Based on the (nitrate + nitrite) contents in Kuala Bubon the water was classified as mesotrophic, while based on phosphate content the water was classified as hypertrophic since the average nutrient content was 1.31 mg L-1 (nitrate+nitrite) and phosphate, 0.12 mg L-. Meanwhile, in Kuala Tadu, the water was also categorized as mesotrophic since nitrate+nitrite was 1.078 mg L-1 and phosphate was 0.22 mg L-1 indicate hypertrophic waters as shown in Table 3. The nitrate+nitrite content in Kuala Bubon was higher than in Kuala Tadu, however, the phosphate content in Kuala Tadu was higher than in Kuala Bubon.
Table 2. The range of water quality measured in situ and ex situ at nipa ecosystem of Aceh Province, Indonesia
|
No. |
Parameter |
Location |
|
|
Kuala Bubon |
Kuala Tadu |
||
|
1 |
Temperature (℃) |
22 - 26 |
23 - 27 |
|
2. |
Salinity (ppt) |
15 - 20 |
14 – 20 |
|
3. |
Turbidity (cm) |
30 - 50 |
30 - 40 |
|
4. |
DO (mg L-1) |
5.4 – 6.0 |
5.6 – 6.0 |
|
5. |
pH |
6.6 – 7.2 |
6.6 – 6.9 |
|
6. |
COD (mg L-1) |
45.05 – 53.25 |
40.96 - 49.15 |
Table 3. The average nitrate + nitrite and phosphate contents in nipa N. fruticans ecosystem at Kuala Bubon and Kuala Tadu, Aceh Province, Indonesia
|
Location |
Parameter |
Average value |
Category |
|
Kuala Bubon |
Nitrate+Nitrite (mg L-1) |
1.31 |
Mesotrophic |
|
Phosphate (mg L-1) |
0.12 |
Eutrophic |
|
|
Kuala Tadu |
Nitrate+Nitrite (mg L-1) |
1.08 |
Mesotrophic |
|
Phosphate (mg L-1) |
0.22 |
Eutrophic |
This study showed that based on the number of species, Baciliariophycea dominated the phytoplankton community in Kuala Bubon, while Dinophyceaceans or Dinoflagellates were dominant in Kuala Tadu. Based on the number of individuals, Amphisolenia bidentate was dominant in Kuala Tadu, while Haramonas sp. was dominant in Kuala Bubon. Haramonas sp. was also found in Kuala Tadu with an almost similar number of individuals. The results were in agreement with Marlian [26] who recorded that Bacillariophycea and Dinophyceae are the two dominant phytoplankton groups in Kueng Cangkoi Estuary, Meulaboh, West Aceh. A similar result was also reported by Sarinda et al. [27] in the waters of Kuala Gigieng, Aceh Besar. This result was also supported by Aryawati et al. [28] who stated that Bacillariophyceae and Dinophyceae were dominant in various coastal and estuary waters in Indonesia. According to Nurdin et al. [3], Bacillariophyceae and Dinophyceacae are distributed widely in coastal and estuary waters because they have a high tolerance and physiological adaptation to environmental changes.
In general, the abundance of phytoplankton in both locations was low, however, two dominant species, including A. bidentata and Haramonas sp. emerged, and Haramonas sp. was dominant in both locations. Haramonas sp. commonly occupy freshwater and acidic swamp waters, and although it is not a harmful alga, red tides caused by its blooming off the coast of Japan have disrupted the production of fish ponds in this area [3]. A similar phenomenon was reported by Irawan et al. [29] stated that algal blooms (HABs) have caused mass mortality of fish in floating net cages at Ringgung Beach, Teluk Lampung, Indonesia. According to several studies, the high abundance of phytoplankton in water has the potential to reduce the aquatic biota population, resulting in the depletion of dissolved oxygen, particularly at night [30, 31]. Therefore, information on species composition and dominance is essential to prevent algae blooms, particularly Dinophyceae (Dinoflagellates), which are toxic and dangerous to aquatic biota [32].
The distribution of phytoplankton is influenced by the physical and chemical parameters of waters [14]. The analysis reveals that the abundance of phytoplankton in Kuala Bubon waters is influenced by turbidity and pH, while in Kuala Tadu it is influenced by COD, turbidity, pH, and dissolved oxygen. Therefore, from the above factors, the turbidity in both locations was high due to the muddy bottom of the waters originating from anthropogenic activities along the river, as well as upstream erosion. During high tide, the mud at the bottom is stirred and rises to the surface, impeding sunlight penetration into the water column. This is in agreement with Costa et al. [33]; Takarina et al. [34] and Pomati et al. [35] who stated that turbidity is closely related to suspended particles in the water including organic matters which limit light penetration and interferes with the photosynthetic activity of phytoplankton, resulting in decreased oxygen production. Therefore, the lower diversity of phytoplankton in Kuala Bubon and Kuala Tadu was probably due to the high nutrient content in both locations, which caused eutrophication.
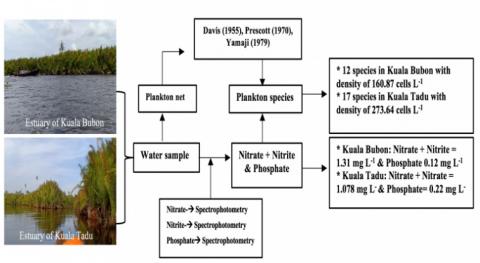
Figure 2. The flow chart of the study
Based nitrate content the waters are classified into 3 categories; oligotrophic waters have nitrate content of 0 - 1 mg L-1, 1 - 5 mg L-1 for mesotrophic, and 5 - 50 mg L-1 for eutrophic waters [36]. While, Hakanson and Bryann cited in Sanusi [37] classified the water based on the phosphate content, oligotrophic (0.000 - 0.020 mg L-1), mesotrophic (0.021 - 0.050 mg L-1), eutrophic (0.051 - 0.100 mg L-1), and hypertrophic (above >0.100 mg L-1). Figure 2 illustrates the workflow and research findings to overview, and provide a general understanding of the research.
According to Rahayu et al. [38] high N and P content indicates the water has been polluted by organic matter that cause eutrophication [3, 39]. The results showed that both locations were classified as mesotrophic to hypertrophic waters, as indicated by the emergence of dominant species in both locations, namely Amphisolnia bidentate and Haramonas sp.
Baho et al. [40] stated that estuary waters have unique characteristics, such as high nutrient load but a low abundance and diversity of plankton. Therefore, a high nutrient content does not necessarily result in a high abundance of phytoplankton, because the high nutrient levels have reduced the quality of estuary waters [3, 35]. High nutrient content disrupts the stability of aquatic ecosystems by triggering the emergence of harmful algae that are tolerant to high nutrients, thereby changing the structure of the plankton community and reducing its abundance [41, 42].
Dinophyceae and Bacillariophyceae were the dominant phytoplankton groups in both locations. In addition, Amphisolenia bidentate and Haramonas sp. were the dominant species in Kuala Tadu. However, Haramonas sp. was also dominant in Kuala Bubon. Based on the nitrate content, the Kuala Bubon and Kuala Tadu were classified as mesotrophic, while based phosphate content both locations were classified as hypertrophic waters with a lack species richness and low abundance of phytoplankton as well as presence of dominant species.
Based on the nitrate and nitrite content, the Kuala Bubon and Kuala Tadu were classified as mesotrophic, while based on phosphate content both locations were classified as hypertrophic waters with a lack of species richness and low abundance of phytoplankton as well as the presence of dominant species. These findings are an early warning that these waters have been contaminated with organic matter. Therefore, it is necessary to plan a better management strategy to anticipate worse conditions.
This study was conducted at the end of the dry season, while the data in the rainy season was not available. It was assumed that may be variations in species composition and abundance in both seasons. Therefore, it is necessary to continue the study in the rainy season to compare the results in both seasons.
The authors would like to thank the Ministry of Education, Culture, Research, and Technology of the Republic of Indonesia for supporting this study by providing a BPPDN scholarship for the first author. The authors would also wish to thank all their colleagues in the Syiah Kuala University and Teuku Umar University laboratories.
[1] Chen, B., Liu, H. (2010). Relationships between phytoplankton growth and cell size in surface oceans: Interactive effects of temperature, nutrients, and grazing. Limnology and Oceanography, 55: 965-972. https://doi.org/10.4319/lo.2010.55.3.0965
[2] Emoyoma, U.O., Numbere, A.O., Woke, G.N. (2019). Impact of Nypa palm (Nypa fruticans) and mangroves forests on plankton community along Andoni River, Nigeria. International Journal Marine Science, 9(4): 36-44. https://doi.org.10.5376/ijms.2019.09.0004
[3] Nurdin, J., Irawan, D., Syandri, H., Nofrita., Rizaldi. (2020). Phytoplankton and the correlation to primary productivity, chlorophyll-a, and nutrients in Lake Maninjau, West Sumatra, Indonesia. AACL Bioflux, 13(3): 1689-1702.
[4] Hastuti, A.W., Pancawati, Y., Surana, I.N. (2018). The Abundance and spatial distribution of plankton communities in Perancak Estuary, Bali. IOP Conferences Series. Earth Environmental Science, 176: 012042. https://doi.org.10.1088/1755-1315/176/1/012042
[5] Kaswadji, R., Hatta, M., Umar, N.A. (2009). Penyusunan model untuk penangkapan berkelanjutan ikan pelagis dengan pendekatan jenjang trofik di Selat Makassar. Jurnal Natur Indonesia, 12(1): 67-74. https://doi.org.10.31258/jnat.12.1.67-74
[6] Mustapha, N., Baharuddin, N., Tan, S.K., Marshall, D.J. (2021). The neritid snails of Brunei Darussalam: Their geographical, ecological and conservation significance. Ecologycal Montenegrina, 42: 45-61. https://doi.org.10.37828/em.202.42.2
[7] Wang, H., Zhao, Y., Zhang, Z., Pang, Y., Liang, D. (2017). Water transparency distribution under varied currents in the largest river-connected lake of China. Water Science and Technology, 75(2): 406-417. https://doi.org.10.2166/wst.2016.534
[8] Smayda, T.J. (2008). Complexity in the eutrophication – harmful algal bloom relationship, with comment on the importance of grazing. Harmful Alga, 8(1): 140-151. https://doi.org.10.1016/j.hal.2008.08.018
[9] Ridhawani, F., Ghalib, M., Nurrachmi, I. (2017). Tingkat kesuburan perairan berdasarkan kelimpahan fitoplankton dan nitrat-fosfat terhadap tingkat kekeruhan muara sungai Rokan Kabupaten Rokan Hilir. Jurnal Perikanan dan Kelautan, 22(2): 10-17. http://dx.doi.org/10.31258/jpk.22.2.10-17
[10] Dzierzbicka-Głowacka, L., Jakacki, J., Janecki, M., Nowicki, A. (2011). variability in the distribution of phytoplankton as affected by changes to the main physical parameters in The Baltic Sea. Oceanologia, 53: 449-470. https://doi.org/10.5697/oc.53-1-TI.449
[11] Saifullah, A.S.M., Hena, A., Kamal, M., Hanafi, M. (2019). Community composition and diversity of phytoplankton in relation to environmental variables and seasonality in a tropical mangrove estuary. Regional Studies in Marine Science, 32: 100826. https://doi.org/10.1016/j.rsma.2019.100826
[12] Muchlisin, Z.A. (2001). Kelimpahan dan keanekaragaman plankton sebagai indicator biologis kerusakan dan pencemaran Sungai Sarah di Kecamatan Lhoknga-Leupung, Kabupaten Aceh Besar. Jurnal Ilmiah MIPA, 3(2): 7-14.
[13] Unbekna, S., Hernawan, U., Setyobudi, E., Wijayanti, L.A.S., Satriyo, T.B., Aryudiawan, C., Setiawan, R.Y. (2020). Plankton abundance and diversity north of Lembata Island, Indonesia. AACL Bioflux, 13: 2355-2364.
[14] Rahayu, D.R.U.S., Anggoro, S., Soeprobowati, T.R. (2020). Plankton community structure and trophic status of Wadaslintang Reservoir, Indonesia. AACL Bioflux, 13: 1138-1151.
[15] Mulyadi, M., Ulqodry, T.Z., Aryawati, R., Isnaini, I., Surbakti, H. (2019). Karakteristik sebaran fitoplankton di perairan muara Sungai Sugihan, Sumatera Selatan. Jurnal Kelautan Tropis, 22(1): 19-26. https://doi.org/10.14710/jkt.v22i1.3178
[16] Jannah, R., Muchlisin, Z.A. (2012). Phytoplankton community in estuary area of Krueng Aceh, Banda Aceh City. Depik, 1(3): 189-195. https://doi.org/10.13170/depik.1.3.121
[17] Davis, C.C. (1955). The Marine and Fresh-Water Plankton. Michigan State University Press, Michigan USA.
[18] Prescott, G.W. (1970). Algae of the western Great Lakes areas. Pub Cranbrook Institute of science Bulletin, 33: 1-496.
[19] Yamaji, I. (1979). Illustrations of the Marine Plankton of Japan. Hoikusha Publishing Co., Japan.
[20] APHA. (1989). Standard methods for examination of water and wastewater. American Public Health Association, Washington DC.
[21] Sandu, M., Lupascu, T., Tarita, A., Goreacioc, T., Turcan, S., Mosanu, E. (2014). Method for nitrate determination in water in the presence of nitrite. Chemistry Journal of Moldova, 9(2): 8-13. https://doi.org/10.19261/cjm.2014.09(2).01
[22] Eisenreich, S.J., Bannerman, R.T., Armstrong, D.E. (1975). A simplified phosphorous analysis technique. Environmental Letters, 9(1): 43-53. https://doi.org/10.1080/00139307509437455
[23] Yamaguchi, H., Hoppenrath, M., Takishita, K., Horiguchi, T. (2008). Haramonas pauciplastida sp. nov. (Raphidophyceae, Heterokontophyta) and phylogenetic analyses of Haramonas species using small subunit ribosomal RNA gene sequences. Phycological Research, 56(2): 127-138. https://doi.org/10.1111/j.1440-1835.2008.00493.x
[24] Daugbjerg, N., Jensen, m.H., Hansen, P.J. (2013). Using Nuclear-encoded LSU and SSU rDNA sequences to identify the eukaryotic endosymbiont in Amphisolenia bidentata (Dinophyceae). Protist, 164: 411-422. https://doi.org/10.1016/j.protis.2012.10.001
[25] Gul, S., Saifullah, S.M. (2007). Genus Amphisolenia stein from North-West Arabian Sea Shelf of Pakistan. Pakistan Journal of Botany, 39(2): 561-576.
[26] Marlian, N. (2017). Analysis of water quality parameters and pollution level status in the Bay Waters of Meulaboh. Proceedings of the International Conference on Integrated Coastal Management and Marine Biotechnology, November, 29-30th 2016, Bogor, Indonesia, pp. 1-7.
[27] Sarinda, F., Dewiyanti, I. (2013). Keragaman fitoplankton di Perairan Estuaria Kuala Gigieng Kabupaten Aceh Besar, Provinsi Aceh. Depik, 2: 20-25. https://doi.org/10.13170/depik.2.1.581
[28] Aryawati, R., Bengen, D.G., Prartono, T., Zulkifli, H. (2017). The abundance of phytoplankton in the coastal waters of South Sumatera. Indonesia Journal of Marine Science, 22(1): 31-39. https://doi.org/10.14710/ik.ijms.22.1.31-39
[29] Irawan, A., Hasani, Q., Yuliyanto, H. (2017). Fenomena harmful algal blooms (HABs) di Pantai Ringgung Teluk Lampung, pengaruhnya dengan tingkat kematian ikan yang dibudidayakan pada karamba jaring apung. Jurnal Penelitian Pertanian Terapan, 15: 48-53. https://doi.org/10.25181/jppt.v15i1.111
[30] Pamukas, N.A., Syafriadiman., Lukistyowati, I., Efriyeldi., Feliatra, F., Mulyadi., Syakti, A.D., Fauzi, M., Windarti. (2020). Types and abundance of plankton in the hybrid tilapia brackish water culture media enriched with mixed booster (plankton, aqua enzyme and amino liquid). AACL Bioflux, 13(2): 1198-1210.
[31] Gurning, L.F.P., Nuraini, R.A.T., Suryono, S. (2020). Kelimpahan fitoplankton penyebab harmful algal bloom di Perairan Desa Bedono, Demak. Journal of Marine Research, 9: 251-260. https://doi.org/10.14710/jmr.v9i3.27483
[32] Fetahi, T., Schagerl, M., Mengistou, S. (2014). Key drivers for phytoplankton composition and biomass in an Ethiopian Highland Lake. Limnologica, 46: 77-83. https://doi.org/10.1016/j.limno.2013.10.007
[33] Costa, C.R., Costa, M.F., Dantas, D.V., Barletta, M. (2018). Interannual and seasonal variations in estuarine water quality. Frontier in Marine Science, 5: 301. https://doi.org/10.3389/fmars.2018.00301
[34] Takarina, N.D., Nurliansyah, W., Wardhana, W. (2019). Relationship between environmental parameters and the plankton community of The Batuhideung Fishing Grounds, Pandeglang, Banten, Indonesia. Biodiversitas, 20: 171-180. https://doi.org/10.13057/biodiv/d200120
[35] Pomati, F., Shurin, J.B., Andersen, K.H., Tellenbach, C., Barton, A.D. (2020). Interacting temperature, nutrients and zooplankton grazing control phytoplankton size-abundance relationships in eight Swiss Lakes. Frontier in Microbiology, 10: 3155. https://doi.org/10.3389/fmicb.2019.03155
[36] Effendi, H. (2003). Telaah kualitas air bagi pengelolaan sumber daya dan lingkungan perairan. Kanisius, Yogyakarta. p. 278.
[37] Sanusi, H. (2006). Kimia laut (Proses fisik kimia dan interaksinya dengan lingkungan). Fakultas Perikanan dan Ilmu Kelauan, Institut Pertanian Bogor, Bogor.
[38] Rahayu, D.R.U.S., Anggoro, S., Soeprobowat, T.R. (2020). Plankton community structure and trophic status of Wadaslintang Reservoir, Indonesia. AACL Bioflux, 3: 1138-1151.
[39] Golmarvi, D., Kapourchali, M.F., Moradi, A.M., Fatemi, M., Nadoshan, R.M. (2017). Influence of physico-chemical factors, zooplankton species. biodiversity and seasonal abundance in Anzali International Wetland, Iran. Open Journal of Marine Science, 7: 91-99. http://dx.doi.org/10.4236/ojms.2017.71007
[40] Baho, D.L., Drakare, S., Johnson, R.K., Allen, C.R., Angeler, D.G. (2016). Is the impact of eutrophication on phytoplankton diversity dependent on lake volume/ecosystem size? Journal of Limnology, 76(1): 199-210. https://doi.org/10.4081/jlimnol.2016.1562
[41] Park, G.S., Marshall, H.G. (2000). Estuarine relationships between zooplankton community structure and trophic gradients. Journal of Plankton Research, 22: 121-135.
[42] Bužančić, M., Gladan, Ž.N., Marasović, I., Kušpilić, G., Grbec, B. (2016). Eutrophication influence on phytoplankton community composition in three bays on The Eastern Adriatic Coast. Oceanologia, 58(4): 302-316. http://dx.doi.org/10.1016/j.oceano.2016.05.003