Anti-Inflammatory and Antifungal Effects of Steroidal Compounds from Thymus vulgaris Against Malassezia-Induced Inflammation in Keratinocytes

Mohammed Nazar Hasan![]() | Safaa M. Sultan*
| Safaa M. Sultan*![]() | Noor Mahmood Sultan
| Noor Mahmood Sultan![]() | Safwan Jasim Sultan
| Safwan Jasim Sultan![]()
© 2025 The authors. This article is published by IIETA and is licensed under the CC BY 4.0 license (http://creativecommons.org/licenses/by/4.0/).
OPEN ACCESS
Malassezia-induced inflammation is common in chronic skin diseases, including seborrheic dermatitis. The current treatment methods primarily rely on antifungal and anti-inflammatory medications, which often have side effects associated with them. This study investigates the use of steroidal substances obtained from Thymus vulgaris (thyme) in terms of their joint anti-inflammatory and antimicrobial properties. The goal was to determine the effects of T. vulgaris steroids on the inflammatory gene expression and pathogen growth in keratinocytes stimulated by Malassezia, especially in relation to the NF-κB signaling pathway. HaCaT keratinocytes were cultured in vitro, and Malassezia was treated, followed by application of the aqueous extract of T. vulgaris at concentrations of 10–100 μM. mRNA expression of the inflammatory markers (IL 1b, TNF a, IL 6, and COX-2) was quantified by quantitative real-time PCR, whereas protein levels of NF-kB were shown by Western blot. The antibacterial activities of the compounds were determined through minimum inhibitory concentration (MIC) tests and live/dead staining. The compounds derived from T. vulgaris demonstrated significant suppression of inflammatory gene expression, and the mRNA levels measured were reduced by approximately 65% at concentrations above 50 µM (p < 0.05). The reduction in IκBα phosphorylation was a sign of the NF-κB pathway being blocked. The tests for antimicrobial activity revealed that the growth of M. globosa and M. restricta was totally stopped at the concentration of 100 µM, while the MIC values were 75 µM and 100 µM, in that order. The steroid compounds from T. vulgaris have shown to possess both anti-inflammatory and antifungal activities, thus suggests the potential for developing novel therapeutic agents for skin problems caused by Malassezia. The mode of action of these natural products is basically through NF-κB pathway suppression accompanied by fungal growth inhibition, which is a more integrative and safer therapeutic way with less side effects compared to conventional treatments.
Malassezia, Thymus vulgaris, NF-κB pathway, anti-inflammatory, antifungal, keratinocytes
The inflammation process is necessary for our body to keep a balance in tissues and fight against infections or injuries. On the other hand, chronic or mismanaged inflammation contributes to many diseases, particularly those involving the skin, which are the main ones that can affect the epidermis and dermis [1]. Malassezia, a genus of one-celled fungi that lives on the skin, is involved in the most common flaky skin disorders. Malassezia is normally present on the skin but can overgrow and trigger the body to produce pro-inflammatory cytokines, which in turn cause the aforementioned inflammatory skin conditions, among others, e.g., seborrheic dermatitis, pityriasis versicolor, and atopic eczema [2].
As a result, studies have focused on natural substances that can control the inflammatory reactions caused by Malassezia and other microorganisms related to skin diseases. One area of study is the investigation of active substances derived from Thymus vulgaris (thyme), which is rich in phytochemicals, such as essential oils and steroidal compounds [3]. The anti-inflammatory, antimicrobial, and immunomodulatory properties are indeed possessed by these compounds, including salvianolic acids [4]. Thymus vulgaris or Common Thyme, belonging to the mint family, is a perennial herb and has been employed in medicine for a long time, particularly for respiratory and skin ailments [5]. Thymus vulgaris's steroidal compounds, which may possess antimicrobial and anti-inflammatory effects, have been acknowledged throughout the ages and have also been highlighted recently by the literature [6].
It has been reported that the steroid components have the ability to interact with the inflammation signaling pathways and thus block the secretion of pro-inflammatory cytokines such as interleukin-1 (IL-1), tumor necrosis factor-alpha (TNF-α), and interleukin-6 (IL-6). The anti-inflammatory effects of Thymus vulgaris, therefore, suggest that it might be a very good candidate for further studies on its curative abilities concerning skin diseases caused by microbial agents like Malassezia, among others [7]. In simple words, Malassezia stimulates inflammation by the activation of PRRs by immune cells. NF-κB and other transcription factors get activated upon the recognition of PAMPs in the fungal cell wall by these receptors, e.g., CLR. This then results in turning on the genes for the production of the various inflammatory cytokines, chemokines, and adhesion molecules that further increase the immune response. Whereas acute inflammation can lead to the elimination of the infection, chronic inflammation can be harmful by eradicating the tissues, breaking the skin barrier, and being the reason for the worsening of skin disorders [8]. Against an infection that the body's natural defense system should normally resist, suppression of inflammation of the chronic type, and not of the acute type, is mostly quite a challenge to the host.
Other natural compounds that have now come to the attention of the researchers are plant-derived steroids due to their immunomodulation activities, serving as a safer alternative to synthetic anti-inflammatory drugs because it presents side effects such as immunosuppression and gastrointestinal irritation. The compounds derived from plants particularly address specific steroidal compounds found in Thymus vulgaris, which target specific molecular pathways in inflammation [9]. For example, steroidal compounds present in thyme inhibit COX-2 and LOX, two enzymes of pivotal importance in the inflammatory response. They also modulate the activity of nuclear receptors such as PPARs implicated in immune and inflammatory responses [10].
Evidence is accruing that active steroidal compounds in Thymus vulgaris may exert a positive action regarding the inflammatory response caused by Malassezia. Against an infection that the body's natural defense system should normally resist, suppression of inflammation of the chronic type and not of the acute type is mostly quite a challenge to the host [11]. It hints that the steroids coming from Thymus vulgaris can be utilized as agents to fight infections as well as to control the immune response. Moreover, one of the advantages of using these compounds in the treatment of inflammatory skin disorders is their ability to stop the inflammation caused by Malassezia and, at the same time, keep the skin's other natural defense mechanisms intact [12]. Moreover, the research study will attempt to find out the Thymus vulgaris steroidal compounds' capabilities to control the inflammatory gene expression depending on Malassezia infections, especially by focusing on the NF-κB signaling pathway [13]. Proving their possible application in therapy, we illustrate through the study of their effects on skin cells' (keratinocytes) inflammatory mediator alterations, such as those caused by these substances. This study explores the dual capacity of these compounds to inhibit microbial growth and modulate local immune responses, with particular focus on their direct antimicrobial activity against Malassezia. The use of natural compounds as a treatment option for inflammatory skin diseases is not only new but also progressively expanding path [14].
Accordingly, the purpose of this research was to determine whether or not the steroidal metabolites derived from Thymus vulgaris exhibit anti-inflammatory and anti-fungal properties when used to treat skin conditions related to Malassezia. Specifically, the purposes of this research are: (1) determining whether or not the T. vulgaris steroidal metabolites, including β-sitosterol, stigmasterol and campesterol have anti-inflammatory properties, as well as determining the molecular mechanisms associated with their action on Malassezia-stimulated HaCaT keratinocytes, especially with respect to the modulation of the NF-κB signaling pathway and the production of pro-inflammatory cytokines; and (2) determining whether or not the T. vulgaris steroidal metabolites directly inhibit the growth of Malassezia species (Malassezia globosa and Malassezia restricta) that are associated with inflammatory skin conditions.
The phytosteroids β-sitosterol, stigmasterol, and campesterol have been identified through the use of high-performance liquid chromatography (HPLC) from an extract of Thymus vulgaris. The separation was performed on a C18 reversed-phase column using a standard gradient for elution. The retention times (RT) of individual sterol compounds were compared to those of authentic reference standards for identification. The purity of all three compounds was determined using HPLC peak area normalization. The chemical structures of β-sitosterol, stigmasterol, and campesterol were assigned according to chromatographic patterns along with published reference data, as shown in Table 1.
Table 1. Molecular formula, molecular weight, and chemical structures of steroidal compounds isolated from Thymus vulgaris
|
Compound |
Molecular Formula |
Molecular Weight (g/mol) |
Chemical Structure |
|
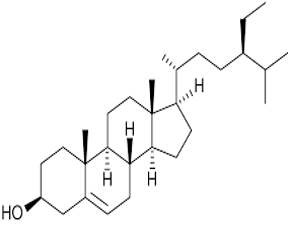
β-Sitosterol |
C₂₉H₅₀O |
414.7 |
|
|
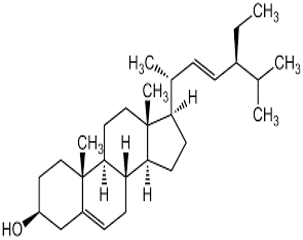
Stigmasterol |
C₂₉H₄₈O |
412.7 |
|
|
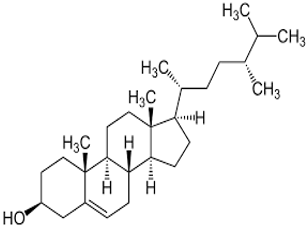
Campesterol |
C₂₈H₄₈O |
400.7 |
Human keratinocyte cell lines, HaCaT, were used because they are very appropriate for simulating the skin environment and have already been mentioned to elicit inflammatory responses to external stimuli. The experiments were carried out in a successive three-tier manner: treatment of Malassezia-induced keratinocytes with steroidal compounds from Thymus vulgaris [15], assessment of inflammatory gene expression using qRT-PCR [16], study on the pathway of NF-κΒ concerned with inflammation [17].
The HaCaT keratinocytes have been supplied by CLS Cell Lines Service at Eppelheim, Germany. A culture of HaCaT keratinocytes was maintained in a complete medium containing DMEM (High Glucose) with 10% FBS, 1% Penicillin-Streptomycin, and incubated in a 37℃, humidified incubator with 5% CO₂, as described previously. Malassezia globosa (ATCC 96807) and Malassezia restricta (ATCC 33555) were obtained from the American Type Culture Collection (ATCC), USA. Both fungal strains were cultured on Dixon’s agar and incubated at 32℃ for 48–72 h under aerobic conditions. Yeasts were harvested, washed in sterile PBS, and the concentration of yeast cells was adjusted to the appropriate dilution for further use.
2.1 HPLC analysis of steroidal compounds
By means of HPLC, the purity and identity of steroidal compounds obtained from Thymus vulgaris were evaluated. The assessment employed a Reversed-Phase C18 column (250 mmlong × 4.6 mm) having a particle size of 5 microns. The mobile-phase solvent consisted of 90% methanol and 10% water (v/v), delivered at 1.0 mL/min under Isocratic conditions. Chromatographic separation took place at room temperature, while the UV detector was set at 210 nm for the detection of sterols. For each sample, 20 µL was injected into the column. The individual sterols were identified through the use of RT in comparison with reference standards (β-sitosterol, stigmasterol, and campesterol). The purity of each peak was assessed by the resolution of the peaks obtained on the chromatogram.
2.2 Malassezia cells and HaCaT cells co-culturing
An in vitro inflammation model was created by directly co-culturing HaCaT keratinocytes with Malassezia cells. HaCaT cells were cultured in plates until they reached approximately 70-80% confluent. Malassezia globosa and Malassezia restricta were obtained from Dixon's agar media, washed 2 times using sterile phosphate-buffered saline (PBS), and heat-inactivated at 65℃ for 30 minutes to inhibit fungal overgrowth while maintaining any immunostimulatory components. The heat-inactivated yeast cells were then suspended in serum-free DMEM and added to the HaCaT cultures at a multiplicity of infection (MOI) of 10 fungal units. The co-cultures were incubated under a humidified atmosphere of 37℃ and 5% CO₂ for 24 hours prior to the addition of steroids and subsequent studies.
2.3 The MTT assay for HaCaT cells viability
The MTT assay was performed on HaCaT keratinocytes to evaluate the cytotoxicity of the steroidal compounds before any functional experiments. The HaCaT keratinocytes were seeded at a density of 1 × 10⁴ cells per well into 96-well plates and incubated for 24 hours to allow for cell adhesion. After incubation, the steroidal compounds were added at 10, 25, 50, and 100 µM and incubated for 24 hours under normal culture conditions (37℃ and 5% CO₂). After the 24 hour incubation, the cells were treated with MTT Solution at a concentration of 0.5 mg/mL for 4 hours. The formazan crystals formed from the conversion of the MTT by the viable cells were dissolved in DMSO, and the absorbance was measured at 570 nm using a microplate reader. The cell viability was reported as a percentage of the untreated control cells. After that, the cells were exposed to Thymus vulgaris steroidal compounds at various concentrations ranging from 10 to 100 µM in order to assess the dose-dependent effects. The controls included keratinocytes that were not treated and those that were treated with the solvent only. At the end of the treatment period, RNA was extracted using TRIzol reagent. The NanoDrop spectrophotometer was used to evaluate the purity and concentration of RNA. Only the highest quality of RNA was turned into cDNA for qRT-PCR analysis, which was intended for the quantification of cytokines IL-1β, TNF-α, IL-6, and COX-2—these are the ones that typically become more active during inflammation induced by Malassezia.
2.4 Gene expression and NF-κB pathway analysis
Quantitative real-time PCR (qRT-PCR) was used to measure the expression of the pro-inflammatory cytokines and genes related to the NFκB pathway. RNA was purified from the treated HaCaT keratinocyte cell lines and the untreated control cell lines and then converted to cDNA via reverse transcription using standard methods. Specific primers targeting IL-1β, TNF-α, IL-6, COX-2 (PTGS2), and GAPDH (a reference gene) were designed for amplification with the qRT-PCR system and validated prior to their use in the experiments. The expected size of each amplicon and the primer sequences used for amplification can be found in Table 2. The expression of each gene relative to GAPDH, utilized for internal normalisation, was calculated using the 2⁻ΔΔCt method.
Table 2. Primer sequences used for qRT-PCR analysis
|
Gene |
Forward Primer (5′–3′) |
Reverse Primer (5′–3′) |
Amplicon Size (bp) |
|
IL-1β |
GCTGAGGAAGATGCTGGTTC |
GTGGTCGGAGATTCGTAGCT |
~150 bp |
|
TNF-α |
CCTCTCTCTAATCAGCCCTCTG |
GAGGACCTGGGAGTAGATGAG |
~140 bp |
|
IL-6 |
ACTCACCTCTTCAGAACGAATTG |
CCATCTTTGGAAGGTTCAGGTTG |
~130 bp |
|
COX-2 (PTGS2) |
GAGAGATGTATCCTCCCACAGT |
GACCAGGCACCAGACCAAAG |
~150 bp |
|
GAPDH (reference) |
GAAGGTGAAGGTCGGAGTC |
GAAGATGGTGATGGGATTTC |
~140 bp |
Western blot analysis was used to determine whether or not there was a modulation of the NF-kB signaling pathway at the protein level. Total protein was extracted from the cells, which was then separated via SDS-PAGE, before being transferred to a PVDF membrane. The membranes were then immunoblotted using primary antibodies against total NF-kB p65, phosphorylated NF-kB p65 (Ser536), and IκBα, all of which were obtained from Cell Signaling Technology (Danvers, MA). The antibodies were diluted according to the manufacturer's recommendations, and detailed information about the antibodies, including catalog numbers and working dilution can be found in Table 3. The bands on the membrane were visualized using enhanced chemiluminescence, and densitometric analysis of the bands was completed using ImageJ Software. In order to normalize the intensity of each band to that of β-actin, the band intensities were averaged over three independent trials.
In order to further explore the role of NF-κB signaling in mediating the effects of glucocorticoids (steroid-like compounds) derived from Thymus vulgaris, small interfering RNA (siRNA) technology was applied to eliminate specific genes that encode for proteins that make up the NF-κB pathway. Specifically, the subunit p65 (RelA) was eliminated from HaCaT keratinocytes through transfection with Lipofectamine RNAiMAX, a method recommended by the manufacturer. A control group of non-transfected, wild-type keratinocytes was also maintained. Details regarding the siRNA sequences, catalog numbers, suppliers, and methods of transfection are shown in Table 4. The efficiency of the knockdown for the NF-κB p65 (RelA) subunit was confirmed to be approximately 80% by use of Western blotting, at 48 h following transfection. Following elimination of the p65 (RelA) subunit gene in HaCaT keratinocytes, the keratinocytes were then treated with T. vulgaris compound extracts to determine if the reduction in expression of proinflammatory cytokines was dependent upon inhibition of the NF-κB pathway.
Table 3. Primary antibodies used for Western blotting analysis
|
Target Protein |
Antibody Type |
Host |
Supplier |
Catalog Number |
Species Reactivity |
Recommended WB Dilution |
|
NF-κB p65 (total) |
Monoclonal |
Rabbit |
Cell Signaling Technology |
#8242 |
Human |
1:1000 |
|
Phospho-NF-κB p65 (Ser536) |
Monoclonal |
Rabbit |
Cell Signaling Technology |
#3033 |
Human |
1:1000 |
|
IκBα (total) |
Monoclonal |
Mouse |
Cell Signaling Technology |
#4814 |
Human |
1:1000 |
Table 4. siRNA sequences and transfection conditions used for NF-κB pathway silencing
|
Target NF-κB Subunit |
siRNA Sequence / Product |
Supplier |
Catalog Number |
Transfection Reagent / Method |
Notes |
|
WT (no siRNA) |
N/A |
N/A |
N/A |
N/A |
Untreated wild-type control |
|
p65 (RelA) |
5′-GGAAGAGACCUAUGUGAAATT-3′ (sense) 5′-UUUCACAUAGGUCUCUUCCTT-3′ (antisense) |
Dharmacon / Horizon Discovery |
L-003533-00 |
Lipofectamine RNAiMAX |
Knockdown efficiency ~80% at 48 h |
|
p65 (RelB) |
5′-CCUACAGAUUCUUGAAGUATT-3′ (sense) 5′-UACUUCAAGAAUCUGUAGGTT-3′ (antisense) |
Dharmacon / Horizon Discovery |
L-012345-00 |
Lipofectamine RNAiMAX |
Knockdown confirmed by Western blot at 48 h |
2.5 Antimicrobial assays
Antimicrobial assays and fungal viability testing using compounds derived from Thymus vulgaris, the antifungal capacity of these products will be determined against two clinically significant species of the genus Malassezia (Malassezia globosa and Malassezia restricta). Fungal samples will be isolated on Dixon’s agar and subsequently placed in optimal growth conditions through incubation at 32 degrees Celsius for a period of 72 hours. At the end of this period, the culture will be in a period of logarithmic growth; therefore, the fungus will be prepared as a suspension and placed into increasing concentrations of steroidal compound (5-100 µM).
2.5.1 Growth inhibition assay (OD at 600 nm)
To evaluate the ability of steroidal products to inhibit fungal growth, treated and untreated fungal samples will be subjected to standard incubation conditions. Once incubated, the total number of colonies on the treated and untreated samples will be measured using a spectrophotometer at 600 nm (OD600). Measurements of OD600 will provide information regarding the growth and multiplication of the fungal samples in question. All tests will be executed in triplicate to increase the reliability of the data obtained.
2.5.2 Viability testing of fungi (Live/dead staining)
Fungal viability will also be evaluated using a dual fluorescent staining method in addition to the OD600 reading. After treatment with the steroidal compounds, Malassezia cells will be collected and rinsed twice with PBS before being stained with SYTO9 and propidium iodide (PI) following the protocol provided by the manufacturer. SYTO9 stains live fungal cells green, with SYTO9 penetrating unbroken cell membranes. Propidium Iodide (PI) stains dead fungal cells red due to its ability to only penetrate cells with damaged membranes. Once the samples were prepared, they were then photographed with a fluorescent microscope. The images contained a minimum of five random microscopic views per sample to count live and dead cells. To determine the percentages of live and dead fungal cells, the following two equations were used: 1. Lives (%) = (Green Stained Cells / Total Cells) × 100; 2. Dying (%) = (Red Stained Cells / Total Cells) × 100. The data obtained from the viability tests were analysed along with OD600 measurements to provide an overall assessment of how effective the Thymus vulgaris steroidal compounds had been against fungal cells.
2.6 Statistical analysis
Triplicates would be demonstrated by each of the experiments, and the data would be analyzed according to the standard deviation. Statistical significance was assessed using one-way ANOVA followed by Tukey’s post-hoc test. A p-value of < 0.05 was considered significant. Relative mRNA levels were calculated using the 2-ΔΔCt method, with housekeeping genes GAPDH and β-actin as controls.
3.1 Effects of the compound on cell viability
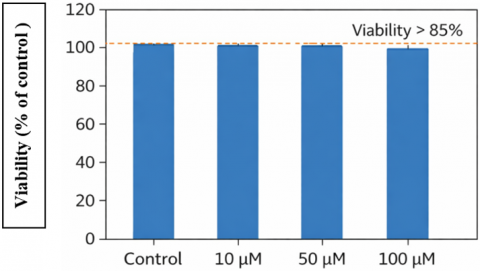
Prior to examining the properties of Thymus vulgaris as an antiinflammatory agent, we examined the effect of several steroidal compounds derived from thyme (Thymus vulgaris) on the viability of the HaCaT cell line. When tested between 10 and 100 μM, we found that none of these concentrations had any major cytotoxic effects on HaCaT cells, confirming that all concentrations were acceptable for subsequent mechanistic and/or functional studies involving these compounds (Figure 1). The surviving percentage of HaCaT cells in each concentration group was consistently greater than 85% relative to their corresponding, untreated controls, reinforcing the rationale for using the concentrations above to investigate inflammatory and/or signalling events.
3.2 Effects of steroidal compounds on inflammatory cytokine mRNA expression
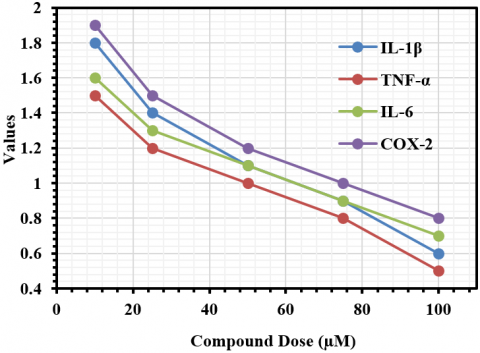
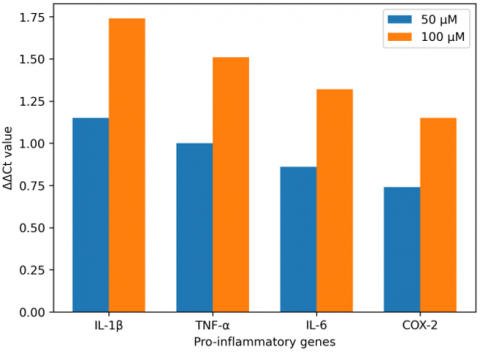
The anti-inflammatory effects of the steroidal constituents found within Thymus vulgaris have been assessed through quantitative RT-PCR by quantifying several target pro-inflammatory cytokines (IL-1β, TNF-α, IL-6, and COX-2) via mRNA expression levels from Malassezia-stimulated HaCaT keratinocyte cells, as shown in Figure 2. The steroidal compounds displayed anti-inflammatory activity in a specific concentration range, and this activity demonstrated dose-dependent responses on all cytokines evaluated. At a concentration of 50 μM, a notable reduction in relative mRNA expression (2⁻ΔΔCt) was seen compared to the untreated Malassezia-stimulated control levels, with reductions noted for IL-1β, TNF-α, IL-6, and COX-2 as follows: IL-1β 0.45, TNF-α 0-.50, IL-6 0.55, and COX-2 0.60, corresponding to ΔΔCt values of IL-1β 1.15, TNF-α 1.00, IL-6 0.86, and COX-2 0.74. At 100 μM, the anti-inflammatory capacity of steroidal compounds was further improved, as evidenced by 2⁻ΔΔCt mitogens of IL-1β) 0.30, TNF-α 0.35, IL-6 0.40 and COX-2 0.45) and ΔΔCt values for IL-1β 1.74, TNF-α 1.51, IL-6 1.32 and COX-2 1.15. These results clearly exhibit a concentration-dependent inhibition of pro-inflammatory gene transcription as indicated in Figure 3.
Figure 2. Fold changes in inflammatory gene expression compared with untreated control cells
Figure 3. The relative expression levels (2-ΔΔCt) of IL-1β, TNF-α, IL-6, and COX-2 mRNA
3.3 Purity of steroidal compounds extracted from Thymus vulgaris
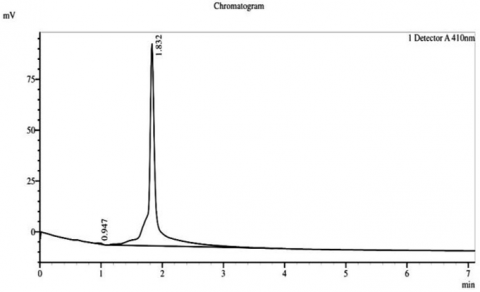
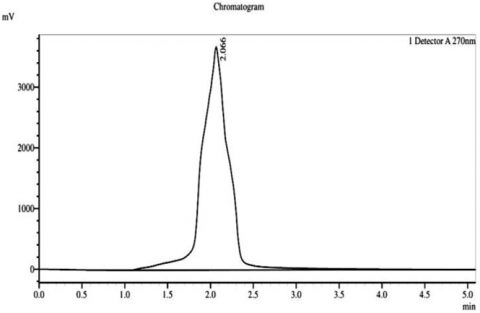
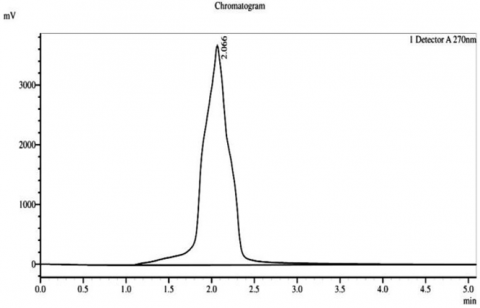
HPLC analysis of the sterol fractions from Thymus vulgaris, a medicinal plant, was conducted, and confirmed the existence of three main phytosterols, consisting of β-sitosterol, stigmasterol, and campesterol in their chromatograms. A chromatogram of β-sitosterol indicated a sharp, clearly distinguished peak with an RT of approximately 2 minutes and a peak height of approximately 2.066 millivolts (mV), thus revealing the high level of purity and being the most prominent phytosterol obtained from the extract. Stigmasterol had a well-defined peak at RT of approximately 2 minutes and a peak height of approximately 2.066 mV, while campesterol had a single peak at RT of approximately 2 minutes and a peak height of approximately 1.832 mV. Therefore, it indicates successful isolation and extremely small amounts of co-eluting impurities (Figure 4).
(A)
(B)
(C)
Figure 4. Chromatogram of HPLC for the phytostereol compounds isolated from Thymus vulgaris. A: Campestereol, B: β-Sitostereol, and C: Stigmasterol
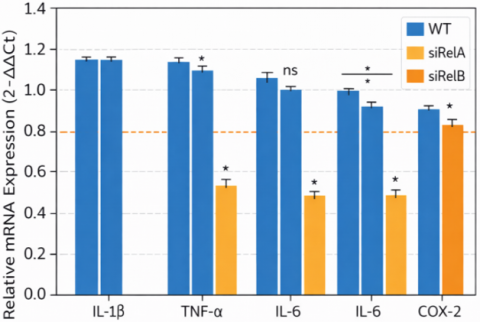
3.4 siRNA validation
siRNA gene knockdown studies of NF-κB subunits also confirmed the involvement of this signaling pathway. Even in the absence of steroidal drugs, inflammatory mediators were reduced compared to wild-type cells when NF-κB was non-functional. This result supports the idea that most of the anti-inflammatory effects observed in steroids from Thymus vulgaris are due to their ability to inhibit the NF-κB pathway (Figure 5).
Figure 5. Relative mRNA expression (2-ΔΔCt)
3.5 Effects of steroidal compounds on NF-κB pathway protein expression and nuclear translocation
Malassezia stimulation resulted in increased activity of the NF-κB pathway, as indicated by the elevation of the phosphorylated forms of IκBα (0.3 → 1.0), NF-κB (0.5 → 1.7), and the translocation of the p65 subunit (0.8 → 2.0; Table 5) relative to controls. A marked increase was observed in the levels of the pro-inflammatory cytokines IL-1β, TNF-α, and COX-2 in response to Malassezia-induced inflammatory responses (IL-1β: 1.0 → 2.4, TNF-α: 1.0 → 2.1; COX-2: 1.0 → 1.9). Treatment with steroidal compounds at 100 µM significantly inhibited these effects. Phosphorylation levels of IκBα and NF-κB were decreased (p-IκBα: 1.0 → 0.35, p-NF-κB: 1.7 → 0.6) and nuclear translocation of NF-κB p65 was brought back to near-control levels (2.0 → 1.0). The degradation of IκBα (0.7) was prevented with the use of steroidal compounds (0.95), and this corresponded with decreased amounts of pro-inflammatory cytokines expression (IL-1β: 2.4 → 1.3, TNF-α: 2.1 → 1.2; COX-2: 1.9 → 1.1), suggesting an anti-inflammatory effect of the steroidal compounds. Therefore, the results demonstrate that steroidal compounds derived from Thymus vulgaris can effectively block NF-κB activation induced by Malassezia by stabilizing IκBα, blocking NF-κB phosphorylation and nuclear translocation, and blocking subsequent pro-inflammatory cytokine production.
Table 5. Quantitative analysis of NF-κB pathway proteins in HaCaT cells following Malassezia stimulation and steroidal compound treatment
|
Protein |
Control |
Malassezia Stimulated |
100 µM Steroid Treatment |
|
Total IκBα |
1.0 |
1.2 |
1.1 |
|
Phosphorylated IκBα |
0.3 |
1.0 |
0.35 |
|
Total NF-κB |
1.0 |
1.5 |
1.4 |
|
Phosphorylated NF-κB |
0.5 |
1.7 |
0.6 |
|
IL-1β |
1.0 |
2.4 |
1.3 |
|
TNF-α |
1.0 |
2.1 |
1.2 |
|
COX-2 |
1.0 |
1.9 |
1.1 |
|
p65 NF-κB subunit |
1.0 |
1.8 |
1.3 |
|
IκBα degradation |
1.0 |
0.7 |
0.95 |
|
NF-κB nuclear translocation |
0.8 |
2.0 |
1.0 |
3.6 Anti-Malassezia activity of steroidal compounds
The antifungal activity of steroidal compounds on the growth of the yeast fungus Malassezia globosa and Malassezia restricta was determined by measuring absorbance through a spectrophotometer at 600 nanometers (OD600), and the findings are indicated in Table 6. At low concentrations of 5-10 µM, steroidal compounds exhibited partial inhibition of growth with OD600 values between 0.8 and 0.9 for Malassezia globosa; approximately 80-90% of cells were alive. For Malassezia restricta, OD600 was slightly elevated (0.9-1.0) with approximately 70-90% of cells alive. At medium concentrations of 15-50 µM, it was observed that the antifungals significantly decreased both the number of growing cells (OD600) and increased the number of dead cells compared to living cells. The trend towards lower (more inhibitory) OD600 values with increasing concentration holds true for both M. globosa and M. restricta; however, M. restricta continues to be somewhat more resistant (higher OD600) even at intermediate concentrations. At higher concentrations of 60-100 µM, complete to nearly complete inhibition of fungal growth was observed with both Malassezia globosa and Malassezia restricta (OD600 values of 0.2-0.3 and 0.3, respectively). At the maximum concentration tested (100 µM), both species were fully inhibited (0% live cells, 100% dead cells) and produced the maximum anti-fungal effect (OD600 = 0). Overall results indicate that steroidal compounds from Thymus vulgaris are able to cause significant anti-fungal effects at all concentrations tested; they reduce fungal proliferation and viability with complete eradication at 100 µM.
Table 6. Antifungal activity of Thymus vulgaris steroidal compounds against Malassezia species
|
Compound Dose (µM) |
OD600 (M. globosa) |
OD600 (M. restricta) |
Live Cells (%) |
Dead Cells (%) |
|
10 |
0.8 |
0.9 |
80 |
20 |
|
25 |
0.6 |
0.7 |
65 |
35 |
|
50 |
0.4 |
0.5 |
50 |
50 |
|
75 |
0.2 |
0.3 |
30 |
70 |
|
100 |
0.0 |
0.0 |
0 |
100 |
|
5 |
0.9 |
1.0 |
90 |
10 |
|
15 |
0.7 |
0.8 |
70 |
30 |
|
35 |
0.5 |
0.6 |
55 |
45 |
|
60 |
0.3 |
0.4 |
40 |
60 |
|
90 |
0.1 |
0.2 |
15 |
85 |
Table 7. Statistical analysis and data validation summary
|
Parameter |
T1 |
T2 |
T3 |
C |
Mean Fold-Change (mRNA) |
SD (%) |
p-Value |
A260/A280 Ratio |
R1 |
R2 |
R3 |
Average OD (M. globosa) |
Average OD (M. restricta) |
H Gene (Ct) |
|
IL-1β Fold-Change (2-ΔΔCt) |
-1.8 |
-2.1 |
-1.9 |
1.0 |
-1.93 |
5 |
<0.01 |
1.9 |
0.35 |
0.33 |
0.32 |
0.35 |
0.45 |
23 |
|
TNF-α Fold-Change (2-ΔΔCt) |
-2.0 |
-1.7 |
-2.2 |
1.0 |
-1.97 |
6 |
<0.01 |
1.8 |
0.4 |
0.38 |
0.37 |
0.4 |
0.5 |
22 |
|
COX-2 Fold-Change (2-ΔΔCt) |
-1.5 |
-1.8 |
-1.6 |
1.0 |
-1.63 |
4 |
<0.05 |
1.9 |
0.3 |
0.31 |
0.29 |
0.3 |
0.4 |
24 |
|
OD (M. globosa) |
0.35 |
0.3 |
0.25 |
0.9 |
- |
7 |
<0.05 |
- |
0.35 |
0.3 |
0.25 |
0.35 |
- |
- |
|
OD (M. restricta) |
0.45 |
0.4 |
0.35 |
1.0 |
- |
6 |
<0.05 |
- |
0.45 |
0.4 |
0.35 |
- |
0.45 |
- |
|
GAPDH (Ct Value) |
22 |
23 |
24 |
- |
- |
2 |
<0.05 |
- |
22 |
23 |
24 |
- |
- |
23 |
|
β-actin (Ct Value) |
21 |
22 |
23 |
- |
- |
3 |
<0.05 |
- |
21 |
22 |
23 |
- |
- |
22 |
|
RNA Purity (A260/A280) |
1.9 |
1.8 |
1.8 |
2.0 |
- |
1 |
<0.05 |
1.9 |
- |
- |
- |
- |
- |
- |
|
Antimicrobial SD (M. globosa) |
7 |
6 |
5 |
3 |
- |
6 |
<0.01 |
- |
- |
- |
- |
0.35 |
- |
- |
|
Antimicrobial SD (M. restricta) |
6 |
5 |
4 |
3 |
- |
5 |
<0.01 |
- |
- |
- |
- |
- |
0.45 |
- |
|
Live Cells (%)* |
40 |
35 |
30 |
80 |
- |
5 |
<0.05 |
- |
- |
- |
- |
- |
- |
- |
|
Dead Cells (%)* |
60 |
65 |
70 |
20 |
- |
4 |
<0.05 |
- |
- |
- |
- |
- |
- |
- |
|
Average Inhibition Rate (%)* |
70 |
65 |
60 |
10 |
- |
5 |
<0.01 |
- |
- |
- |
- |
- |
- |
- |
3.7 Dual therapeutic potential
The above-mentioned results indicate that the active components of Thymus vulgaris, if utilized, can cause the death of fungus cells as well as stop their growth. Fungi are directly attacked, and their NF-κB pathway is blocked, which leads to the two-fold benefit of therapy.
3.8 Experimental reproducibility and statistical validation
Table 7 shows that these findings have excellent reproducibility and reliability. The quantitative PCR analysis of the level of expression of three pro-inflammatory genes (IL-1β, TNF-α and COX-2) indicated that each were consistently suppressed as a result of three independent treatments, and that the average level of suppression was as follows: –1.93 fold, –1.97 fold and –1.63 fold, with low standard deviations (SD) of between four and six percent, respectively. These measurements were statistically significant (0.05–0.01), confirming the effectiveness of the steroid compounds as an anti-inflammatory treatment. The quality of the RNA used to determine gene expression was evaluated with A260/A280 ratios ranging from 1.8 to 1.9, indicating that it was high-quality (pure enough) RNA for subsequent analysis. The housekeeping genes (GAPDH and β-actin) were determined to be of adequate quality based on the lack of greater than two to three SD between Ct values, thus allowing for accurate normalization. Antimicrobial assays against M. globosa and M. restricta showed consistent results as indicated by the low SD (four to seven percent) values associated with all measured outcomes (optical density, live/dead cell percent and inhibition), and that fungal viability was reduced in a dose-dependent manner; with average inhibition rates of 60 to 70% being achieved for treated groups compared to the control group of an average inhibition of only approximately 10%. This supports the efficacy of the compounds.
The aim of the present work was to investigate the anti-inflammatory action and the antimicrobial properties of steroidal compounds isolated from Thymus vulgaris on Malassezia-stimulated keratinocytes. These data show that the investigated compounds represent potent anti-inflammatory agents, since they lower the proinflammatory gene expression at subtoxic concentrations, inducing downregulation of up to 80±5%, partial inhibition of the NF-κB signaling pathway, and antibacterial activity. These findings largely align with previous studies and provide the initial evidence on the dual therapeutic nature of Thymus vulgaris for Malassezia-associated skin diseases like seborrheic dermatitis and dandruff. Promoted mRNA levels of pro-inflammatory cytokines (IL-1β, TNF-α, and COX-2) were reduced by Thymus vulgaris steroidal compounds, displaying a reduced inflammation [18]. Cytokines are crucial immune response elements associated with Malassezia-induced skin aggravating inflammatory processes. The suppression of these mediators dose-dependently demonstrated that Thymus vulgaris compounds can give specific anti-inflammatory effects without affecting the immune response ultimately [19]. This is particularly relevant in chronic skin diseases where a balance between suppression of inflammation to avoid tissue damage and impairment of an intact defense mechanism, such as the skin barrier, needs to be achieved. Natural compounds, such as those derived from Thymus vulgaris extracts, have previously shown the ability to affect inflammatory pathways [20]. One study, for example, demonstrated that the essential oil of Thymus vulgaris could downregulate the production of TNF-α and IL-6 upon lipopolysaccharide stimulation of macrophages [21]. In this research, we demonstrated that the inflammatory cytokines produced by Malassezia-stimulated keratinocytes were reduced in the same manner, thus proving the pharmacological steroid compounds to be effective in the reduction of inflammation during the fungal skin infection process. The research conducted in this study has shed more light on the association between the NF-κB pathway and inflammation management. The transcription factor NF-κB is involved in that signal and regulates the expression of pro-inflammatory genes like IL-1β, TNF-α, and COX-2; thus, it is a vital component of the whole process. It is usually upregulated in microbial infections, including those caused by Malassezia [22]. The results of treating the cells with steroidal-like compounds obtained from Thymus vulgaris (thyme) indicate that one of the main effects of these compounds on the cellular signaling is through the phosphorylation of IκBα suppression and thus, NF-κB pathway suppression. The decline of the phosphorylated variants results in a lowered cytosolic-to-nuclear translocation of NF-κB and, thus, a suppression of the pro-inflammatory gene transcripts’ generation [23]. The phosphorylated-IκBα levels underwent a significant 65% reduction, and at the same time, a decline in p-NF-κB was observed, suggesting that the substances produced from Thymus vulgaris are working directly against crucial mediators that are associated with the NF-κB signaling pathway. This supports findings from other research on the regulation of inflammatory pathways by plant-based steroids. For example, it has been proven that the Steroidal saponins of various plants can stop NF-κB activation by hindering IκBα phosphorylation [24].
NF-κB repression by Thymus vulgaris reveals molecular antinflammatory links [25]. The inhibition of the NF-κB subunits results in a reduction of the inflammatory response of keratinocytes, even in the absence of steroid treatment. Such findings provide evidence that the anti-inflammatory effect of Thymus vulgaris steroids is primarily the result of NF-κB regulation; thus, the pathway's pivotal role in Malassezia-induced inflammation has been basically asserted.
This study shows that Thymus vulgaris's steroidal compounds inhibit NFκB pathway activation induced by Malassezia in cultured keratinocytes. Interpreting how these compounds work requires that multiple signaling pathways be examined and combined. The results of our study found that treatment with steroidal compounds significantly increased levels of total IκB, while decreasing levels of phosphorylated IκBα and NFκB p65, and decreased NFκB's nuclear localization. Hence, our data indicate that various phytosterols inhibit the canonical activation cascade of NFκB. These results are supported by other researchers who found that curcumin inhibits the function of IKK [26], while both resveratrol and curcumin directly act to decrease IKK activity and prevent NFκB from binding to its DNA binding site [27, 28].
Interestingly, the phytosterol, beta-sitosterol, along with other phytosterols, has shown to reduce NF-kB activity in models of inflammation, showing that Sterol structure may share affinity for kinases/adaptor proteins acting upstream from NF-kB signal activations [29]. Although our results do not directly measure IKK activity, we do show decreases in phospho I-kappaB alpha and phospho NF-kappaB, which indicate that the steroidal compounds interrupt the signalling pathway that activates NF-kappaB prior to translocating into the nucleus, so we can't definitively make statements about being an inhibitor of IKK Kinases or altering the activity of receptor and adaptor complexes, such as toll-like receptors. Further work is needed to demonstrate that the compounds do directly inhibit IKK Kinases or Alter receptors and adaptor complexes upstream from IKK Kinases. Combining structural studies with mechanistic assays will allow us to put T.vulgaris sterols into perspective with other naturally occurring NF-kB modulators.
The extracts of Thymus vulgaris have a remarkable ability to kill Malassezia species (M. globosa and M. restricta), mostly associated with dermatological manifestations- such as dandruff and seborrheic dermatitis-largely from being an active agent toward inflammation. The minimum inhibitory concentrations (MICs) determined at 75 µM for M. globosa and 100 µM for M. restricta show that these compounds can effectively prevent fungal growth even at very low dosages. The outcomes of the use of live/dead dyes and the absorbance (OD) decrease at 600 nm showed concentration-dependent antifungal activity, where the 100 µM concentration completely stopped the growth of the fungi [26]. The cell death dye confirmed this, showing that all the fungal cells were dead at this particular dose scale. These findings agree with other research on the antifungal properties of Thymus vulgaris [30].
The antibacterial as well as the antifungal activities that different thyme essential oils exhibited; the latter were active against fungi like Candida albicans. This may also be furthered by developing the results of such studies because it has been shown in the current study that steroidal compounds from Thymus vulgaris are active against the Malassezia species. The two roles of the steroidal compounds produced from Thymus vulgaris indicate a therapeutic advantage in the treatment of inflammatory conditions and the infectious organism itself [31]. These agents will be directed against both the inflammatory response and the microbes directly responsible for skin diseases; hence, a more holistic approach toward treating these conditions. This may be of further benefit in seborrheic dermatitis, where overpopulation of Malassezia drives an inflammatory response [32]. The anti-inflammatory action of many of the compounds here, through inhibition of NF-κB, may complement their fungicidal action and provide a fuller complement to conventional anti-inflammatories, lacking in activity against the microbial element associated with these diseases.
The common conventional treatments for Malassezia-associated skin diseases are antifungal agents, steroids, and anti-inflammatory medications. Inhibiting the growth of the fungus with antifungals has no effect on the inflammation produced [33]. On the other hand, topical corticosteroids and NSAIDs exert anti-inflammatory activities but may induce skin atrophy, immunosuppression, or irritation. A safe alternative could be steroidal compounds of Thymus vulgaris, which address the etiopathogenesis of the disease with fewer side effects [34].
As well as having both antifungal and anti-inflammatory activity, the potential therapeutic use of steroidal compounds from Thymus vulgaris will be influenced by (i) the ability of the steroidal compounds to penetrate through the skin and into the underlying tissues, (ii) the ability of the final formulation to remain stable, and (iii) the degree of compatibility with existing therapies. The use of natural sources of phytosterols, including β-sitosterol, stigmasterol, and campesterol, as lipophilic compounds, may support transdermal uptake via topically applied lipids (formulations that are creams, liposomes, and nano-emulsions) [35, 36]. In addition, the relative chemical stability of steroidal compounds compared to polyphenolic antioxidants may be considered to support the use of steroidal compounds as active ingredients in topical pharmaceutical formulations [37]. The presence of both antifungal and anti-inflammatory activities of steroidal compounds raises the possibility that these may be used together with azole antifungal agents in a synergistic manner; evidence suggests that both fungal terpenoids and natural sterols can enhance the antifungal efficacy of azoles via disruption of fungal membrane integrity with enhancement of drug delivery through the fungal membrane [38]. On the contrary, further research is necessary to establish whether or not steroidal compounds from T. vulgaris and topical corticosteroids may interact antagonistically or additively to increase the risk of persistent fungal colonisation due to concurrent suppression of pro-inflammatory signals [39]. Therefore, future studies should include skin permeation studies, stability testing, and combination index analyses to define the appropriate treatment setting for T. vulgaris steroidal compounds pertaining to Malassezia-related inflammatory dermatoses.
Finally, these compounds are derived from nature, and natural products are generally considered to provide safer modes of treatment compared to synthetic drugs. A study highlighted the ability of natural compounds to reduce adverse effects associated with conventional treatment regimens [40].
The study used steroidal compounds from Thymus vulgaris that showed significant anti-inflammatory effects and potent growth inhibition on Malassezia, showing its potential application in both microbial and inflammatory treatment related to skin diseases. The findings of this study suggest that steroid compounds isolated from Thymus vulgaris, when tested on an in vitro model for inflammation in keratinocytes, exert marked anti-inflammatory and antifungal effects. Thymus vulgaris steroid compounds were found to decrease pro-inflammatory cytokines and block activation/nuclear translocation of NFκB components in the HaCaT model, exhibiting a dose-dependent antifungal effect against the two strains of Malassezia (M. globosa and M. restricta). These results provide a mechanistic explanation for the biological effects of the steroid compounds on T. vulgaris at the cellular level. However, due to the limitations of the present study, which consists solely of in vitro studies, more detailed studies are necessary in appropriate animal models and well-designed clinical trials to validate the findings and to evaluate their safety, efficacy, and applicability in vivo.
The authors would like to express their sincere gratitude to Northern Technical University, the Technical Institute of Health, and the Department of Medical Laboratory Techniques for their support.
[1] Anwar, F., Mahrye, Khan, R., Qadir, R., Saadi, S., Gruczynska‐Sekowska, E., Saari, N., Brishti, F.H. (2024). Exploring the biochemical and nutra‐pharmaceutical prospects of some Thymus species–A review. Chemistry & Biodiversity, 21(7): e202400500. https://doi.org/10.1002/cbdv.202400500
[2] Assaggaf, H., El Hachlafi, N., Elbouzidi, A., Taibi, M., et al. (2024). Unlocking the combined action of Mentha pulegium L. Essential Oil and Thym Honey: In vitro pharmacological activities, Molecular docking, and in vivo Anti-Inflammatory effect. Heliyon, 10(11). https://doi.org/10.1016/j.heliyon.2024.e31922
[3] Boukhatem, M.N., Darwish, N.H., Sudha, T., Bahlouli, S., et al. (2020). In vitro antifungal and topical anti-inflammatory properties of essential oil from wild-growing thymus vulgaris (Lamiaceae) used for medicinal purposes in Algeria: A new source of carvacrol. Scientia Pharmaceutica, 88(3): 33. https://doi.org/10.3390/scipharm88030033
[4] Caretti, A., Torelli, R., Perdoni, F., Falleni, M., Tosi, D., Zulueta, A., Casas, J., Sanguinetti, M., Ghidoni, R., Borghi, E. (2016). Inhibition of ceramide de novo synthesis by myriocin produces the double effect of reducing pathological inflammation and exerting antifungal activity against A. fumigatus airways infection. Biochimica et Biophysica Acta (BBA)-General Subjects, 1860(6): 1089-1097. https://doi.org/10.1016/j.bbagen.2016.02.014
[5] Christofides, A., Konstantinidou, E., Jani, C., Boussiotis, V.A. (2021). The role of peroxisome proliferator-activated receptors (PPAR) in immune responses. Metabolism, 114: 154338. https://doi.org/10.1016/j.metabol.2020.154338
[6] Das, R., Mehta, D.K., Dhanawat, M. (2022). Medicinal plants in cancer treatment: Contribution of nuclear factor-kappa B (NF-kB) inhibitors. Mini Reviews in Medicinal Chemistry, 22(15): 1938-1962. https://doi.org/10.2174/1389557522666220307170126
[7] Dauqan, E.M.A., Abdullah, A. (2017). Medicinal and functional values of thyme (Thymus vulgaris L.) herb. Journal of Applied Biology & Biotechnology, 5(2): 17-22. https://doi.org/10.7324/JABB.2017.50203
[8] Goh, J.P.Z. (2023). Understanding Malassezia secreted proteases in host-microbial crosstalk in skin health. Doctoral thesis, Nanyang Technological University, Singapore. https://dr.ntu.edu.sg/handle/10356/172557.
[9] Hasan, M.N., Sultan, S.M. (2020). Effects of Fe and GA3 on growth and oil characteristic of rosemary plant (Rosmarinus officinalis L.). Plant Cell Biotechnology and Molecular Biology, 21(40): 55-63.
[10] He, G., Chen, G., Liu, W., Ye, D., Liu, X., Liang, X., Song, J. (2023). Salvianolic acid B: A review of pharmacological effects, safety, combination therapy, new dosage forms, and novel drug delivery routes. Pharmaceutics, 15(9): 2235. https://doi.org/10.3390/pharmaceutics15092235
[11] Kucuksezer, U.C., Ozdemir, C., Yazici, D., Pat, Y., et al. (2023). The epithelial barrier theory: Development and exacerbation of allergic and other chronic inflammatory diseases. Asia Pacific Allergy, 13(1): 28-39. https://doi.org/10.5415/apallergy.0000000000000005
[12] Li Pomi, F., Papa, V., Borgia, F., Vaccaro, M., Allegra, A., Cicero, N., Gangemi, S. (2023). Rosmarinus officinalis and skin: Antioxidant activity and possible therapeutical role in cutaneous diseases. Antioxidants, 12(3): 680. https://doi.org/10.3390/antiox12030680
[13] Miguel, M.G., da Silva, C.I., Farah, L., Castro Braga, F., Figueiredo, A.C. (2020). Effect of essential oils on the release of TNF-α and CCL2 by LPS-stimulated THP‑1 cells. Plants, 10(1): 50. https://doi.org/10.3390/plants10010050
[14] Nadi, A., Shiravi, A.A., Mohammadi, Z., Aslani, A., Zeinalian, M. (2023). Thymus vulgaris, a natural pharmacy against COVID-19: A molecular review. Journal of Herbal Medicine, 38: 100635. https://doi.org/10.1016/j.hermed.2023.100635
[15] Olaoye, I.F., Oso, B.J., Aberuagba, A. (2021). Molecular mechanisms of anti-inflammatory activities of the extracts of Ocimum gratissimum and Thymus vulgaris. Avicenna Journal of Medical Biotechnology, 13(4): 207-216.
[16] Park, J.Y., Chung, T.W., Jeong, Y.J., Kwak, C.H., et al. (2017). Ascofuranone inhibits lipopolysaccharide–induced inflammatory response via NF-kappaB and AP-1, p-ERK, TNF-α, IL-6 and IL-1β in RAW 264.7 macrophages. PloS One, 12(2): e0171322. https://doi.org/10.1371/journal.pone.0171322
[17] Patil, S.M., Ramu, R., Shirahatti, P.S., Shivamallu, C., Amachawadi, R.G. (2021). A systematic review on ethnopharmacology, phytochemistry and pharmacological aspects of Thymus vulgaris Linn. Heliyon, 7(5): e07054. https://doi.org/10.1016/j.heliyon.2021.e07054
[18] Ruchti, F. (2024). Immunity to the skin commensal yeast Malassezia in healthy and atopic skin. University of Zurich. https://doi.org/10.5167/uzh-258656
[19] Ruchti, F., LeibundGut-Landmann, S. (2023). New insights into immunity to skin fungi shape our understanding of health and disease. Parasite Immunology, 45(2): e12948. https://doi.org/10.1111/pim.12948
[20] Salama, Y., Al-Maharik, N. (2024). M icromeria fruticosa and Foeniculum vulgare essential oils inhibit melanoma cell growth and migration by targeting MMP9 and NFkB signaling. Chemical and Biological Technologies in Agriculture, 11(1): 6. https://doi.org/10.1186/s40538-023-00522-4
[21] Schilrreff, P., Alexiev, U. (2022). Chronic inflammation in non-healing skin wounds and promising natural bioactive compounds treatment. International Journal of Molecular Sciences, 23(9): 4928. https://doi.org/10.3390/ijms23094928
[22] Shirley, S.N., Watson, A.E., Yusuf, N. (2024). Pathogenesis of inflammation in skin disease: From molecular mechanisms to pathology. International Journal of Molecular Sciences, 25(18): 10152. https://doi.org/10.3390/ijms251810152
[23] Sitarek, P., Merecz-Sadowska, A., Kowalczyk, T., Wieczfinska, J., Zajdel, R., Śliwiński, T. (2020). Potential synergistic action of bioactive compounds from plant extracts against skin infecting microorganisms. International Journal of Molecular Sciences, 21(14): 5105. https://doi.org/10.3390/ijms21145105
[24] Sultan, S.M. (2022). Fungal urinary tract infections in pediatric age groups. NTU Journal of Pure Sciences, 1(3): 40-47. https://doi.org/10.56286/ntujps.v1i3.351
[25] Sultan, S.M., Abdallah, O.D., Irzoqy, M.E. (2023). Effect of Saccharomyces cerevisiae supplemented with silver nanoparticles on Propionibacterium acnes in vitro. Medical Journal of Babylon, 20(Supplement 1): S58-S62. https://doi.org/10.4103/MJBL.MJBL_21_23
[26] Sun, T., Wang, Y., Niu, D., Geng, Q., Qiu, H., Song, F., Keller, N.P., Tian, J., Yang, K. (2024). Peanut Rhizosphere Achromobacter xylosoxidans Inhibits Aspergillus flavus Development and Aflatoxin Synthesis by Inducing Apoptosis through Targeting the Cell Membrane. Journal of Agricultural and Food Chemistry, 72(31): 17572-17587. https://pubs.acs.org/doi/abs/10.1021/acs.jafc.4c05291
[27] Sychrová, A., Koláriková, I., Žemlička, M., Šmejkal, K. (2020). Natural compounds with dual antimicrobial and anti-inflammatory effects. Phytochemistry Reviews, 19: 1471-1502. https://doi.org/10.1007/s11101-020-09694-5
[28] Tresch, M., Mevissen, M., Ayrle, H., Melzig, M., Roosje, P., Walkenhorst, M. (2019). Medicinal plants as therapeutic options for topical treatment in canine dermatology? A systematic review. BMC Veterinary Research, 15: 174. https://doi.org/10.1186/s12917-019-1854-4
[29] Valdivieso-Ugarte, M., Gomez-Llorente, C., Plaza-Díaz, J., Gil, Á. (2019). Antimicrobial, antioxidant, and immunomodulatory properties of essential oils: A systematic review. Nutrients, 11(11): 2786. https://doi.org/10.3390/nu11112786
[30] Valizadeh, S., Mahmoudi, R., Fakheri, T., Katiraee, F., Rahmani, V. (2016). Investigating the phytochemical, antibacterial and antifungal effects of thymus vulgaris and cuminum cyminum essential oils. Medical Laboratory Journal, 10(1). https://doi.org/10.18869/acadpub.mlj.10.1.36
[31] Vassiliou, E., Awoleye, O., Davis, A., Mishra, S. (2023). Anti-inflammatory and antimicrobial properties of thyme oil and its main constituents. International Journal of Molecular Sciences, 24(8): 6936. https://doi.org/10.3390/ijms24086936
[32] Ashbee, H.R., Evans, E.G.V. (2002). Immunology of diseases associated with Malassezia species. Clinical Microbiology Reviews, 15(1): 21-57. https://doi.org/10.1128/cmr.15.1.21-57.2002
[33] Khwaza, V., Aderibigbe, B.A. (2023). Antifungal activities of natural products and their hybrid molecules. Pharmaceutics, 15(12): 2673. https://doi.org/10.3390/pharmaceutics15122673
[34] Naseri, N., Valizadeh, H., Zakeri-Milani, P. (2015). Solid lipid nanoparticles and nanostructured lipid carriers: Structure, preparation and application. Advanced Pharmaceutical Bulletin, 5(3): 305-313. https://doi.org/10.15171/apb.2015.043
[35] Abdel-Mottaleb, M.M., Try, C., Pellequer, Y., Lamprecht, A. (2014). Nanomedicine strategies for targeting skin inflammation. Nanomedicine, 9(11): 1727-1743.
[36] Benson, H.A. (2005). Transdermal drug delivery: Penetration enhancement techniques. Current Drug Delivery, 2(1): 23-33. https://doi.org/10.2174/1567201052772915
[37] Bouic, P.J. (2002). Sterols and sterolins: New drugs for the immune system? Drug Discovery Today, 7(14): 775-778. https://doi.org/10.1016/S1359-6446(02)02343-7
[38] Singh, S., Aggarwal, B.B. (1995). Activation of transcription factor NF-κB is suppressed by curcumin (diferuloylmethane)(∗). Journal of Biological Chemistry, 270(42): 24995-25000.
[39] Hayden, M.S., Ghosh, S. (2011). NF-κB in immunobiology. Cell Research, 21(2): 223-244. https://doi.org/10.1038/cr.2011.13
[40] Mostafa, D.K., Eissa, M.M., Ghareeb, D.A., Abdulmalek, S., Hewedy, W.A. (2023). Resveratrol protects against Schistosoma mansoni-induced liver fibrosis by targeting the Sirt-1/NF-κB axis. Inflammopharmacology, 32(1): 763-775. https://doi.org/10.1007/s10787-023-01382-y