Molecular Epidemiology and Antimicrobial Resistance of ESBL- and Carbapenemase-Producing Uropathogenic E. coli and K. pneumoniae in Community-Acquired UTIs in Baghdad, Iraq

Yasameen Hasan Ali![]()
© 2025 The author. This article is published by IIETA and is licensed under the CC BY 4.0 license (http://creativecommons.org/licenses/by/4.0/).
OPEN ACCESS
Urinary tract infections (UTIs) are common community-acquired infections caused by Escherichia coli and Klebsiella pneumoniae. This study examined the patterns of antimicrobial resistance and the genetic factors responsible for extended-spectrum β-lactamase production in E. coli and K. pneumoniae isolates collected from UTI patients. This study analyzed 42 E. coli and K. pneumoniae isolates from female UTI patients in Baghdad. Phenotypic and genotypic methods assessed antibiotic resistance, plasmid types, clonal relationships, and horizontal gene transfer using polymerase chain reaction (PCR), multilocus sequence typing, Etest, disk diffusion, conjugation, electroporation, and sequencing techniques. The results revealed high resistance rates to multiple antibiotics, including ciprofloxacin (92.31%), gentamicin (76.92%), amoxicillin-clavulanic acid (92.31%), ceftazidime (87.18%), and cefotaxime (76.92%). Imipenem showed the highest sensitivity (76.92%). Molecular analysis identified blaCTX-M-15 in K. pneumoniae ST15 and E. coli ST131 clones, while blaKPC-2 was detected in carbapenem-resistant E. coli. Plasmid profiling revealed IncF-type plasmids as key vectors for resistance gene dissemination. The fingerprinting techniques confirmed clonal transmission, with identical isolates detected across hospitals, suggesting nosocomial spread. Notably, a patient persistently colonized with KPC-2-producing E. coli acted as a potential reservoir. The research findings underscore that the clonal spread of ST131 and KPC-2 carriage represents significant nosocomial threats, warranting urgent containment measures.
Escherichia coli, Klebsiella pneumoniae, ESBL, ST131, plasmid-mediated resistance, carbapenem resistance, plasmid dissemination
Urinary tract infections (UTIs) rank among the most prevalent community-acquired infections, substantially impacting morbidity and mortality rates [1, 2]. They can affect the lower and upper urinary tract, ranging from asymptomatic bacteriuria to severe conditions, such as pyelonephritis and septic shock [3]. UTIs pose a major health concern in both hospital and community settings, particularly among older adults with comorbidities. The primary bacterial pathogens, including Escherichia coli and Klebsiella pneumoniae, are frequently associated with complicated and recurrent UTIs [4]. According to recent research conducted in Iraq, the incidence of UTIs ranges from 15% to 25%, with a greater frequency among women. More than 50% of isolates are multidrug-resistant (MDR) bacteria, especially ESBL-producing E. coli and K. pneumoniae, underscoring the significant concerns about antibiotic resistance [5, 6]. The misuse of antibiotics is a major factor in the rise of multidrug-resistant (MDR) E. coli that produce extended-spectrum β-lactamases (ESBLs) [7]. In Iraq [8], the resistance of uropathogenic E. coli to third-generation cephalosporins increased from 12.1% in 2010 to 14.1%, with ESBL-producing E. coli accounting for 40% of community-acquired UTIs. The number of Enterobacteriaceae that produce ESBL is increasing worldwide. In many regions of the world, blaCTX-M-15 and blaCTX-M-14 have displaced other genotypes, according to research that documented temporal patterns in blaCTX-M epidemiology. Horizontal gene transfer (HGT) of plasmids, successful E. coli clones, ESBLs in food animals, the natural environment, human migration, and access to basic sanitation are some of the explanations for these alterations [9]. Empirical treatment typically involves oral antibiotics, with nitrofurantoin recommended as a first-line option. ESBLs hydrolyze β-lactam antibiotics, rendering penicillin, cephalosporins, and monobactams ineffective. Resistance genes, which are frequently found on plasmids, aid in the horizontal gene transfer that spreads resistance. Plasmids such as IncF are crucial in disseminating blaCTX-M and blaKPC-2 genes. Clones like E. coli ST131 and K. pneumoniae ST15 have been key drivers of the global ESBL spread [8, 10].
Phenotypic characterization was performed using disk diffusion and E-test methods to assess antibiotic susceptibility and determine minimum inhibitory concentrations (MICs) [11, 12]. Genotypic analysis involves polymerase chain reaction (PCR) and sequencing to detect resistance genes, plasmid types, and mobile genetic elements [13, 14]. Molecular typing techniques, including multilocus sequence typing (MLST) and PCR fingerprinting, were used to determine clonal relationships [14, 15]. Horizontal gene transfer was evaluated through conjugation and electroporation experiments [16, 17]. This study analyzed MDR bacterial isolates from UTI patients in three Baghdad hospitals (Al-Yarmouk Teaching Hospital, Al-Shaab General Hospital, and Baghdad Medical City) between March 2023 and April 2024. The objective was to investigate the genetic mechanisms underlying antibiotic resistance and identify potential reservoirs facilitating bacterial persistence and transmission.
The patterns of antibiotic resistance and the genetic factors responsible for the synthesis of extended-spectrum β-lactamase in K. pneumoniae and E. coli isolates obtained from UTI patients were investigated in this study. Identifying key resistance genes and their plasmid associations emphasizes the need to implement enhanced surveillance, infection control measures, and antimicrobial stewardship programs to mitigate the spread of resistance.
2.1 Study population
This cross-sectional study evaluated E. coli and K. pneumoniae isolates from female urinary tract infection (UTI) patients for antibiotic resistance across three hospitals in Baghdad, Iraq, between March 2023 and April 2024. Forty-two [9] bacterial isolates were recovered exclusively from women with an average age of 40.32 years.
2.2 Collection of urine samples
Urine samples were collected from all the participants. To minimize contamination, urine samples were collected using the midstream clean-catch technique [18]. For hospitalized patients, catheterized urine samples were obtained under sterile conditions. Additionally, blood samples were collected from patients suspected of having systemic infections to assess bacteremia [2, 19].
2.3 Inclusion and exclusion criteria
The study included female patients diagnosed with UTIs based on positive urine cultures and clinical symptoms. Patients were excluded if they had non-bacterial infections or had received antibiotics within 72 hours before sampling.
2.4 Antibiotic susceptibility testing
Antibiotic susceptibility testing was conducted using the Etest and disk diffusion (Kirby-Bauer) methods to determine MICs [12, 20]. Bacterial isolates were cultured on Drigalski agar and incubated at 37℃ for 24 hours. Suspensions were adjusted to 0.5 McFarland (1×10⁶ CFU/mL) and inoculated onto Mueller-Hinton (MH) agar. The Kirby-Bauer disk diffusion method was used to evaluate antibiotic susceptibility, and the chosen antibiotics are listed in Table 1. The results were interpreted according to EUCAST (2013) guidelines [21]. Inhibitors such as boronic acid (20 mg/mL) and cloxacillin (10 mg/mL) were used to detect group A carbapenemases and AmpC β-lactamases. MICs were determined via the Etest method using CTX, CAZ, CFEP, IPM, and MEM strips. Combination strips (CTX/CTXL, CAZ/CAZL, CFEP/FEPL) confirmed ESBL production, while IPM/IPI strips detected metallo-β-lactamases. Conjugation and electroporation experiments were performed to evaluate horizontal gene transfer, providing insight into the dissemination of resistance genes among UTI-causing bacteria in Iraqi hospitals.
Table 1. Antibiotics used in susceptibility testing
|
Antibiotics |
Abbreviation |
Disc Conc. (μg) |
|
Ciprofloxacin |
CIP |
5 |
|
Gentamicin |
GEN |
10 |
|
Cefoxitin |
FOX |
30 |
|
Cefotaxime |
CTX |
30 |
|
Ceftazidime |
CAZ |
30 |
|
Amoxicillin/ Clavulanic Acid |
AMC |
20/10 |
|
Cefepime |
CFEP |
30 |
|
Imipenem |
IPM |
10 |
|
Meropenem |
MEM |
10 |
|
Doripenem |
DOR |
10 |
|
Ertapenem |
ETP |
10 |
2.5 Molecular typing and genetic analysis
The genetic analysis was used to identify resistance genes and plasmid types through polymerase chain reaction (PCR) and sequencing. Clonal relationships were assessed using molecular typing methods, such as MLST and PCR fingerprinting [22]. Total DNA was extracted using the boiling-centrifugation method. Genomic characterization was performed using M13-PCR and BOX-PCR fingerprinting, followed by MLST of E. coli and K. pneumoniae isolates [21, 23], targeting seven housekeeping genes. Sequences were analyzed using reference databases. Resistance genes, including blaKPC,blaCTX, blaTEM, blaSHV, blaDHA, blaCMY, blaOXA-48, blaOXA-58, and blaCEFT, were screened via PCR. In carbapenem-resistant K. pneumoniae, porin genes (OmpK35, OmpK36) were analyzed to assess permeability alterations.
2.6 Genetic environment and plasmid analysis
The blaKPC-2 gene’s genetic surroundings, including ISKpn7, ISKpn6, tnpA, and tnpR, were examined. ISEcp1 and IS903, commonly linked to blaCTX-M, were also studied. Class I integrons were identified using primers targeting conserved regions flanking resistance gene cassettes [24]. Plasmid incompatibility groups were determined via replicon typing using multiplex and simplex PCR reactions. Plasmid DNA was extracted using the Kado and Liu [25] method, visualized on agarose gels, and analyzed for β-lactamase genes.
2.7 Genetic transfer methods
Horizontal gene transfer was assessed through conjugation, transformation, and electroporation. E. coli J53 (sodium azide-resistant) served as the recipient strain [26, 27]. Liquid and solid medium conjugation methods were used, followed by transconjugant selection on MH agar with sodium azide and cefotaxime [28]. Competent E. coli JM109 cells were transformed with purified plasmid DNA via electroporation and selected on MH agar. PCR amplification and sequencing confirmed the genetic transfer, with sequences analyzed through BLAST and Clustal Omega [29].
3.1 Distribution of resistance genes and plasmid replicons in E. coli and K. pneumoniae isolates
The main genetic results of E. coli and K. pneumoniae isolates, organized by shared traits, are presented in Table 2. All E. coli isolates with identical genetic environments and plasmid types carried the blaKPC-2 gene, indicating a probable clonal origin. In contrast, K. pneumoniae isolates possessed the blaCTX-M-15 gene and were split into representative groups depending on shared insertion sequences (e.g., ISKpn7, ISKpn8, ISEcp1, and IS903) [30]. Also, Table 2 illustrates how β-lactamase genes spread in hospital settings through plasmid-mediated transmission and clonal expansion.
Table 2. Plasmid-mediated β-lactamase resistance and mobile genetic elements in E. coli and K. pneumoniae from tertiary healthcare facilities
|
Sample Number |
Hospital/Unit |
Resistance Genes (β-lactamases) |
Genetic Environment |
Plasmid Origin of Replication |
KPC |
CTX |
SHV |
TEM |
ISKpn6 |
ISKpn7 |
ISKpn8 |
ISEcp1 |
IS903 |
|
E. coli Isolate |
|||||||||||||
|
1 |
AY-TH (Nephrology) |
blaKPC-2 |
Identical to isolates 2, 3, 49 |
IncFIA, IncFIB, IncF |
+ |
ND |
ND |
ND |
+ |
- |
- |
- |
- |
|
2 |
AY-TH (Emergency) |
blaKPC-2 |
Identical to isolates 1, 3, 49 |
IncFIA, IncFIB, IncF |
+ |
ND |
ND |
ND |
+ |
- |
- |
- |
- |
|
3 |
AY-TH (Emergency) |
blaKPC-2 |
Identical to isolates 1, 2, 49 |
IncFIA, IncFIB, IncF |
+ |
ND |
ND |
ND |
+ |
- |
- |
- |
- |
|
49 |
ASGH (general ward) |
blaKPC-2 |
Identical to isolates 1, 2, 3 |
IncFIA, IncFIB, IncF |
+ |
ND |
ND |
ND |
+ |
- |
- |
- |
- |
|
K. pneumoniae Isolate |
|||||||||||||
|
35 |
Three hospital units |
blaCTX-M-15 |
Representative of a 13-isolate group |
IncF |
- |
+ |
ND |
ND |
- |
+ |
+ |
+ |
+ |
|
40 |
Three hospital units |
blaCTX-M-15 |
Representative of a 13-isolate group |
IncF |
- |
+ |
ND |
ND |
- |
+ |
+ |
+ |
+ |
|
7 |
Three hospital units |
blaCTX-M-15 |
Representative of a 4-isolate group |
IncF |
- |
+ |
ND |
ND |
- |
+ |
+ |
+ |
+ |
|
17 |
Three hospital units |
blaCTX-M-15 |
Representative of a 4-isolate group |
IncF |
- |
+ |
ND |
ND |
- |
+ |
+ |
+ |
+ |
|
29 |
Three hospital units |
blaCTX-M-15 |
Representative of a 4-isolate group |
IncF |
- |
+ |
ND |
ND |
- |
+ |
+ |
+ |
+ |
|
43 |
Three hospital units |
blaCTX-M-15 |
Representative of a 4-isolate group |
IncF |
- |
+ |
ND |
ND |
- |
+ |
+ |
+ |
+ |
The isolates were grouped based on their common characteristics; +: Present; -: Absent; ND: Not detected; KPC: Klebsiella pneumoniae carbapenemase; CTX: Cefotaxime; SHV: Sulfhydryl variable β-lactamase; TEM: Temoniera β-lactamase; ISKpn6: Insertion Sequence Klebsiella pneumoniae 6; ISKpn7: Insertion Sequence Klebsiella pneumoniae 7; ISKpn8: Insertion Sequence Klebsiella pneumoniae 8; ISEcp1: Insertion Sequence for Escherichia coli promoter 1; IS903: Insertion Sequence 903.
3.2 Selection and characteristics of isolates
Seven K. pneumoniae isolates were selected based on their antibiotic resistance profiles. Four isolates were resistant to cefotaxime (CTX) and ceftazidime (CAZ), while three isolates demonstrated intermediate susceptibility or were susceptible to CTX but resistant to CAZ and CFEP. These isolates were selected primarily due to their susceptibility or intermediate resistance to IPM and MEM, alongside resistance to DOR and ETP. Five Enterobacter isolates were selected: two were responsive to carbapenems but resistant to CAZ and CEFT, and three were resistant to cephalosporins (CTX, CAZ, and ceftazidime (CEFT)) and carbapenems (IPM and ETP).
3.3 Antibiotic susceptibility
Antibiograms were performed using the disk diffusion method on Mueller-Hinton (MH) agar to evaluate the resistance profiles of the 42 Enterobacteriaceae isolates. The isolates were obtained from various hospital services, with detailed resistance data provided. The results for key antibiotic categories are presented in Figure 1. A high proportion of the isolates showed resistance to ciprofloxacin (92.31%) and gentamicin (76.92%), as well as to β-lactam antibiotics including amoxicillin-clavulanic acid (92.31%), ceftazidime (87.18%), and cefotaxime (76.92%). Imipenem demonstrated the highest sensitivity percentage at 76.92% among antibiotics. Most imipenem-sensitive isolates are K. pneumoniae strains that resist cephalosporins CEFZ and CTX. The imipenem-resistant isolates consist of four E. coli strains exhibiting resistance to the cephalosporins CEFZ and CTX. The results revealed three primary categories of multidrug-resistant bacteria. The first included four E. coli isolates resistant to the carbapenem imipenem (IPM) and the cephalosporins cefotaxime (CTX) and cefazolin (CEFZ). The second group comprised 19 K. pneumoniae isolates resistant to CTX and CEFZ but susceptible to carbapenems, suggesting the presence of extended-spectrum β-lactamases (ESBLs), particularly CTX-M types. The graph was constructed using the values recorded from the antibiograms of 39 of the 42 isolates under study, since the number of antibiotics tested for E. cloacae isolates 5, 8, and 14 was not sufficient to construct the graph.
Figure 1. Percentage of isolates demonstrating resistance, intermediate levels of resistance, and sensitivity to the test antibiotics
Gentamicin (GNTM), ciprofloxacin (CFX), amoxicillin with clavulanic acid (AMXCA), cefoxitin (CEFT), ceftazidime (CEFZ), cefotaxime (CTX), and imipenem (IPM).
blue symbol refers to the percentage of resistant isolates to antibiotics
red symbol refers to the percentage of intermediate levels of resistance to antibiotics
green symbol refers to the percentage of sensitive isolates to antibiotics
3.4 β-lactamase inhibitory activity
Inhibitor assays were conducted with 10 μL of boronic acid or cloxacillin added to antibiotic discs to detect group A carbapenemases and AmpC β-lactamases. A positive result was defined as an increase in the inhibition zone of more than 5 mm. Seven E. coli isolates were initially included in the study; however, two were excluded due to the absence of cephalosporin resistance. Four isolates (1, 2, 3, and 49) exhibited resistance to all tested antibiotics, including cephalosporins and carbapenems. However, isolate 37 was sensitive to carbapenems but resistant to CTX and CAZ, indicating the development of an ESBL of the CTX-M type.
3.5 Carbapenemase and ESBL confirmation
Boronic acid significantly increased inhibition zones for MEM, ETP, and DOR in isolates 1, 2, 3, and 49, confirming K. pneumoniae carbapenemase (KPC) production. However, cloxacillin and imipenem + EDTA (IPM/IPI) did not affect inhibition zones, ruling out AmpC β-lactamases and metallo-β-lactamases. MIC was determined using Etest strips. For isolate 37, CTX, CTX/CTXL, CAZ, CAZ/CAZL, CFEP, and CFEP/FEPL were tested. For isolates 1, 2, 3, and 49, imipenem (IPM), meropenem (MEM), and imipenem + EDTA (IPM/IPI) were used. MIC values ranged from 8 to 24 mg/L for IPM and MEM (Figure 2), confirming carbapenemase production. There were no significant changes in susceptibility with the use of IPM/IPI strips, effectively ruling out metallo-β-lactamase production. Isolate 37 exhibited minimum inhibitory concentrations (MICs) greater than 256 mg/L for cefotaxime (CFX), cefazolin (CFZ), and cefepime (CEFP), supporting the classification of this isolate as an extended-spectrum β-lactamase (ESBL) producer. The use of CFX/CFXL, CFZ/CFZL, and CEFP/CEFPL combination strips enables the identification of ESBL-producing isolates by comparing results against established reference thresholds: CFX MICs ≥ 0.5 mg/L with a CFX/CFXL ratio ≥ 8; CFZ MICs ≥ 1 mg/L with a CFZ/CFZL ratio ≥ 8; or CEFP MICs ≥ 0.25 mg/L with a CEFP/CEFPL ratio ≥ 8. In the case of isolate 37, the results obtained with the CFZ/CFZL and CEFP/CEFPL strips met these criteria, thereby confirming the presence of ESBL activity.
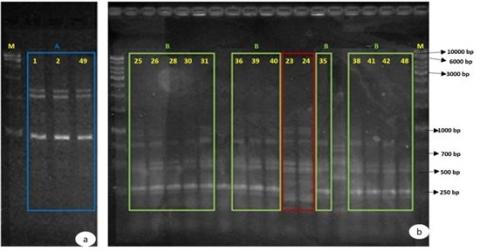
Figure 2. Electrophoretic profiles generated by M13 and BOX fingerprinting molecular typing methods using 2% agarose gel
A: Profile of E. coli isolates; B: Profile of K. pneumoniae isolates; M: GeneRulerTM 1kb DNA ladder molecular weight marker.
Tests were conducted on K. pneumoniae isolates using inhibitors to assess β-lactamase activity. Cloxacillin was added to discs containing ceftazidime (CAZ), amoxicillin/clavulanic acid (AMX/CA), cefotaxime (CTX), cefepime (CFEP), and imipenem (IPM). The lack of alterations in inhibitory zones suggested that AmpC β-lactamases were not produced by these isolates. Another inhibitor, boronic acid, was tested with imipenem (IPM), meropenem (MEM), doripenem (DOR), and ertapenem (ETP). No significant changes were observed in inhibition zones, except in a group of seven isolates where an increased inhibition zone in the DOR + boronic acid disc suggested the presence of a carbapenemase.
The Etest technique was used to verify the existence of ESBLs. With ranges of 8 mg/L to >256 mg/L for cefotaxime (CTX), 12 mg/L to >256 mg/L for ceftazidime (CAZ), and 24 mg/L to >256 mg/L for CFEP, the minimum inhibitory concentrations (MIC) for all three medications were noticeably high. Resistance to these medicines was validated by these high MIC values. Furthermore, when clavulanic acid was diffused into CTXL, CAZL, or FEPL zones, the inhibition ellipse on CTX, CAZ, or CFEP strips showed a "ghost zone" or distortion, which suggested the existence of ESBLs. This trait was most noticeable in K. pneumoniae isolate 24, which was sensitive to carbapenems but resistant to cephalosporins. The molecular typing techniques analyzed the genetic relationships between isolates, including M13-PCR, BOX-PCR fingerprinting, and MLST. In E. coli isolates, M13-PCR fingerprinting established a genetic link between isolates 1, 2, and 49, while BOX-PCR fingerprinting demonstrated similarities between isolate 3 and the other E. coli isolates. These analyses confirmed a close genetic relationship among isolates 1, 2, 3, 49, and 37 from Yarmouk Teaching Hospital. Notably, isolates 1 and 2 were collected from the same patient at different times, one from the nephrology unit and another three months later from the emergency room, while isolates 3, 49, and 37 were obtained from different biological samples of unrelated patients across multiple hospital units.
Among the 19 selected K. pneumoniae isolates, two distinct genetic profiles, B and C, were identified. Profile B included 13 isolates from different patients across three hospitals. In contrast, Profile C consisted of isolates collected from a single patient within a four-day interval at two hospitals, namely Al-Shaab General Hospital and Baghdad Medical City. In contrast, molecular typing of Enterobacter isolates and the seven selected K. pneumoniae isolates did not provide sufficient resolution to establish a clear genetic relationship. Further details on sample origins, hospital sources, and electrophoretic profiles are provided in Table 3. The MLST technique was performed to characterize the bacterial isolates further and confirm their clonal relationships. This method involves amplifying internal fragments of seven housekeeping genes, which are constitutive and species-specific [31]. Based on nucleotide variations, each isolate was assigned an allelic profile that, when analyzed against a reference database, allows for identifying different sequence types (STs) through homology with previously described profiles. Klebsiella pneumoniae isolates 35 and 40 (Profile B), isolate 24 (Profile C), and E. coli isolates 1, 3, 49, and 37 (Profile A) were all subjected to MLST. Additionally, it was performed on K. pneumoniae isolates 7, 17, and 29, which exhibited resistance to cephalosporins and intermediate resistance to carbapenems in antibiograms. In E. coli isolates 1, 3, 49, and 37, the globally disseminated clone ST131 [32] was identified, which is recognized for its role in the spread of the CTX-M-15 enzyme. The allelic profile of these isolates was determined by sequencing seven housekeeping genes [31]: adk, fumC, gyrB, icd, mdh, purA, and recA. The detailed allelic profiles of these isolates are provided in Table 4.
Table 3. Strain identification, sample type, hospital service, unit of origin, and electrophoretic profiles of the studied isolates
|
Isolate |
Isolate No. |
Sample |
Hospital Service Section |
Hospital |
Electrophoretic Profile |
|
Escherichia coli |
1 |
Urine |
Nephrology |
AY-TH |
A |
|
2 |
Urine |
General Urgency |
ASGH |
A |
|
|
49 |
Urine |
Kidney Transplant |
AY-TH |
A |
|
|
37 |
Urine |
Nephrology |
AY-TH |
A |
|
|
Klebsiella pneumoniae |
25 |
Urine |
UCI Internment |
BMC |
B |
|
26 |
Urine |
Kidney Transplant |
AY-TH |
B |
|
|
30 |
Urine |
UCIC Orthopedics |
ASGH |
B |
|
|
31 |
Urine |
Oncology |
ASGH |
B |
|
|
36 |
Urine |
Urology |
BMC |
B |
|
|
39 |
Urine |
UCI Internment |
BMC |
B |
|
|
40 |
Urine |
Internment Medicine IV |
ASGH |
B |
|
|
35 |
Urine |
Urology |
BMC |
B |
|
|
38 |
Urine |
UCI Internment Cardiothoracic Surgery |
AY-TH |
B |
|
|
48 |
Urine |
Hospitalization for Cardiothoracic Surgery |
AY-TH |
B |
|
|
17 |
Urine |
Internment Infectiology |
BMC |
X |
|
|
29 |
Urine |
Hematology |
ASGH |
X |
|
|
43 |
Urine |
Kidney Transplant |
AY-TH |
X |
|
|
46 |
Urine |
Hematology |
ASGH |
X |
AY-TH: Al-Yarmouk Teaching Hospital; ASGH: Al-Shaab General Hospital; BMC: Baghdad Medical City; A: found in E. coli isolates; B: found in K. pneumoniae isolates with one grouping, and X: found in K. pneumoniae isolates with a distinct genetic profile.
Table 4. Allelic profile of E. coli isolates
|
Isolate |
Sequence Type |
adk |
fumC |
gyrB |
icd |
mdh |
purA |
recA |
|
1 |
ST131 |
P |
P |
P |
P |
P |
P |
P |
|
3 |
ST131 |
P |
P |
P |
P |
P |
P |
P |
|
49 |
ST131 |
P |
P |
P |
P |
P |
P |
P |
|
37 |
ST131 |
P |
P |
P |
P |
P |
P |
P |
P: Present (indicates detecting the characteristic nucleotide sequence for each housekeeping gene in ST131 isolates, and ST131 is a globally disseminated clone frequently associated with the spread of the CTX-M-15 extended-spectrum β-lactamase enzyme).
To properly describe the bacterial isolates and validate their clonal connection, the MLST methodology was applied to the most representative profiles that were generated using the earlier techniques. Following analysis of the profiles in a database, the various sequence types may be identified based on their similarity with the previously mentioned profiles. This technique was used for E. coli isolates 1, 3, 49, and 37 (Profile A) and for K. pneumoniae isolates 35 and 40 (Profile B) and 24 (Profile C). It was also applied to K. pneumoniae isolates 7, 17, and 29, showing a resistance profile to cephalosporins and intermediate resistance to carbapenems in the antibiograms. In E. coli isolates 1, 3, 49, and 37, the globally disseminated clone ST131 was identified, which is known for its role in spreading the CTX-M-15 enzyme (Table 4). The allelic profile of these isolates was determined by analyzing the nucleotide sequences of seven housekeeping genes, including adk, fumC, gyrB, icd, mdh, purA, and recA. The detailed allelic profiles of these isolates are presented in Table 4.
ST15, a clone that has recently been linked to the spread of CTX-M enzymes in isolates of K. pneumoniae, was found in isolates 35, 40, 7, 17, and 29 [24]. A distinct ST was found in isolate 24, which is in line with the profiles derived using the BOX fingerprinting typing technique. The closest ST to the one discovered in this isolate is ST20, whose nucleotide sequences of the genes under study vary from isolate 24 only in the mdh gene, as shown in Table 5. The ST detected in this isolate is not mentioned in the database that was utilized. Considering the nucleotide sequences of the housekeeping genes gapA, infB, mdh, pgi, phoE, rpoB, and tonB, Table 5 shows the allelic profile for K. pneumoniae isolates 35, 40, 24, 7, 17, and 29 [33].
Table 5. Allelic profile of K. pneumoniae isolates
|
Isolate |
ST Type |
gapA |
infB |
mdh |
pgi |
phoE |
rpoB |
tonB |
Profile in Database |
|
35 |
ST15 |
"I" |
"I" |
"I" |
"I" |
"I" |
"I" |
"I" |
"I" |
|
40 |
ST15 |
"I" |
"I" |
"I" |
"I" |
"I" |
"I" |
"I" |
"I" |
|
7 |
ST15 |
"I" |
"I" |
"I" |
"I" |
"I" |
"I" |
"I" |
"I" |
|
17 |
ST15 |
"I" |
"I" |
"I" |
"I" |
"I" |
"I" |
"I" |
"I" |
|
29 |
ST15 |
"I" |
"I" |
"I" |
"I" |
"I" |
"I" |
"I" |
"I" |
|
24 |
NE(Closest:ST20) |
"I" |
"I" |
"I" |
"I" |
"I" |
"I" |
"I" |
"I" |
“I”: identical allelic sequences matching those found in ST15 or the nearest sequence type; Variant: a sequence difference, as seen in the mdh gene of isolate 24; Profile in Database: reflects whether the full allelic profile corresponds to an existing entry in the MLST database.
3.6 Genetic determinants of resistance
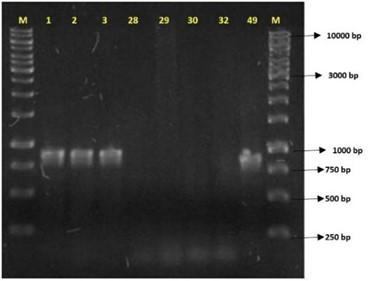
PCR analysis was performed to detect resistance genes (Figure 3), including the carbapenemases blaKPC, blaOXA-48, and blaOXA-58, and ESBLs, blaCTX, blaTEM, blaSHV, blaDHA, blaCMY, and blaCEFT. Enterobacter isolates 28, 29, 30, and 32 tested negative for blaCTX and blaKPC and were excluded from further study. Their resistance may be due to alternative mechanisms, such as porin modifications or efflux pump activity. In E. coli isolates 1, 2, 3, and 49, blaKPC amplification correlated with carbapenem resistance and increased inhibition zones in boronic acid disk tests. Sequencing confirmed the blaKPC-2 variant, the most globally widespread KPC enzyme, primarily found in K. pneumoniae. The blaTEM gene was also detected, identifying the TEM-1 β-lactamase [34]. No amplification was observed for blaCTX, blaOXA-48, blaOXA-58, or blaSHV. The E. coli ST131 clone, known for spreading the CTX-M-15 enzyme, was first reported carrying blaKPC-2 in Ireland [35]. Its presence in this highly transmissible lineage raises concerns about the rapid dissemination of carbapenem resistance.
Figure 3. PCR products related to amplifying the blaKPC gene of 871 bp (base pairs) in a 1% agarose gel
M: GeneRulerTM 1kb DNA ladder molecular weight marker.
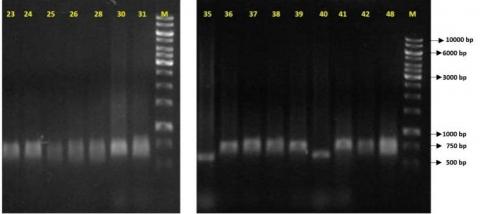
PCR amplification detected the blaCTX gene in E. coli isolate 37 and K. pneumoniae isolates 25, 26, 28, 30, 31, 36, 39, 40, 35, 24, 38, 41, 42, and 48, all of which were resistant to CTX and CEFZ but susceptible to carbapenems. Sequencing confirmed the blaCTX-M-15 variant in isolates 37, 36, 40, 35, and 24, a globally prevalent enzyme conferring resistance to third- and fourth-generation cephalosporins, such as CTX, CEFZ, and cefepime (CFEP). The blaTEM gene was also identified, and sequencing of E. coli isolate 37 and K. pneumoniae isolates 40 and 24 confirmed the blaTEM-1 variant, responsible for resistance to first-generation penicillins and cephalosporins. The blaSHV gene was absent in E. coli isolate 37 but detected in K. pneumoniae isolates 35, 40, and 24. Sequencing revealed blaSHV-28 in isolates 35 and 40 and blaSHV-12 in isolate 24. The SHV-12 enzyme demonstrated enhanced hydrolytic activity against cephalosporins, particularly CEFZ and CTX, while SHV-28 preferentially hydrolyzes CTX over CEFZ, reflecting the resistance patterns observed in isolates 35 and 40. The blaOXA-48 gene was absent in all E. coli and K. pneumoniae isolates. Among seven K. pneumoniae isolates (7, 17, 29, 43, 33, 46, and 47) with susceptibility or intermediate resistance to IPM and MEM but resistance to doripenem (DOR) and ertapenem (ETP), no amplification of blaKPC, blaOXA-48, blaDHA, blaCMY, or blaCEFT was observed. However, isolates 7, 17, 29, and 43, resistant to CTX and CEFZ, tested positive for blaCTX, blaTEM, and blaSHV (Figure 4). Sequencing of isolate 17 confirmed blaCTX-M-15, blaTEM-1, and blaSHV-1. In isolates 33, 46, and 47, which showed intermediate or susceptible levels to CTX but resistance to ceftazidime (CEFT) and CFEP, only the blaSHV-1 gene was identified, conferring resistance to first-generation penicillins and cephalosporins.
Figure 4. PCR products related to amplifying the 544 bp blaCTX gene in a 1% agarose gel
M: GeneRulerTM 1kb DNA ladder molecular weight marker.
Table 6 outlines β-lactamase genes, porin presence, electrophoretic profiles, and sequence types of E. coli and K. pneumoniae isolates. Most E. coli belonged to ST131 harboring blaKPC-2, while K. pneumoniae mainly carried blaCTX-M-15 and belonged to ST15, indicating clonal and gene-specific resistance patterns.
Resistance to β-lactam antibiotics can result from altered outer membrane permeability due to reduced expression or absence of porin-encoding genes. Since the seven K. pneumoniae isolates (7, 17, 29, 33, 43, 46, and 47) with susceptibility or intermediate resistance to IPM and MEM but resistance to DOR and ETP tested negative for blaKPC, the presence of the OmpK35 and OmpK36 porins was investigated. Most non-ESBL-producing K. pneumoniae express both porins, while ESBL-producing isolates commonly express only OmpK36. The absence of both porins has been linked to β-lactam resistance. Recent studies suggest that OmpK35 is associated with ceftazidime susceptibility, whereas OmpK36 expression correlates with cefotaxime susceptibility. Loss of OmpK36 is linked to increased resistance to cefoxitin and carbapenems in β-lactamase-producing strains. The OmpK35 gene was found in all isolates, while OmpK36 was detected in only five (7, 17, 29, 43, and 46). OmpK36 was absent in isolates 33 and 47, which might account for their increased vulnerability to ceftazidime. The absence of OmpK36 may contribute to increased carbapenem resistance. Further analysis is needed to determine porin expression levels [36], as the presence of a gene does not guarantee its expression. Based on these findings, the following isolates were selected for further study: E. coli ST131 isolates 1, 2, 3, and 49 producing KPC-2 carbapenemase; E. coli ST131 isolate 37 producing ESBL CTX-M-15 (Profile A); K. pneumoniae isolate 24 with a distinct sequence type (Profile C); K. pneumoniae isolates 35 and 40 with ST15 (Profile B); and K. pneumoniae isolates 7, 17, 29, and 43 with distinct electrophoretic profiles (X) but sharing ST15, all producing ESBLCTX-M-15.
Table 6. Primary resistance genes identified in the studied isolates and their corresponding hospitals of origin
|
E. coli Isolate Sample No. |
β-Lactamase Genes |
Porins (OmpK35/OmpK36) |
Electrophoretic Profile |
ST Type |
|
1 |
blakpc-2 |
NA |
NA |
ST131 |
|
2 |
blakpc-2 |
NA |
NA |
ST131 |
|
3 |
blakpc-2 |
NA |
NA |
ST131 |
|
37 |
blactx-M-15 |
NA |
A |
ST131 |
|
49 |
blakpc-2 |
NA |
NA |
ST131 |
|
K. pneumoniae |
|
|
|
|
|
7 |
blactx-M-15 |
Present/Present |
X |
ST15 |
|
17 |
blactx-M-15 , blatem-1, blashv-1 |
Present/Present |
X |
ST15 |
|
24 |
Ctx-m-15 |
NA |
C |
Distinct ST |
|
29 |
blactx-M-15 |
Present/Present |
X |
ST15 |
|
33 |
Blashv-1 |
Present/Present |
NA |
NA |
|
35 |
blactx-M-15 |
NA |
B |
ST15 |
|
40 |
blactx-M-15 |
NA |
B |
ST15 |
|
43 |
blactx-M-15 |
Present/Present |
X |
ST15 |
|
46 |
blashv-1 |
Present/Present |
NA |
NA |
|
47 |
blashv-1 |
Present/Absent |
NA |
NA |
NA: Not applicable or not available; ST: Sequence type based on molecular typing; EP: Electrophoretic profile (distinct molecular profiles of isolates).
Table 7. Primers designed in the laboratory and PCR conditions used to determine the genetic environment of the blaKPC-2 gene
|
Primer |
Sequence (5' → 3') |
Initial Denaturation |
No. of Cycles |
Cycle Conditions |
Final Extension |
|
ISKpn8 |
F:GGTGCCTCACTGATTAAGCA |
95℃, 11 min |
35 |
95℃, 46 sec |
73℃, 11 min |
|
R:CAGTTCACCGACCGCTTCA |
57℃, 46 sec |
||||
|
73℃, 2 min |
|||||
|
ISKpn8T |
F:TCATGCCATCCGTAAGATGC |
95℃, 11 min |
35 |
95℃, 46 sec |
73℃, 11 min |
|
R:CCAACTCCTTCAGCAACAAA |
57℃, 46 sec |
||||
|
73℃, 2 min |
|||||
|
KPCT |
F:CATCCGAAAGTGGCAAAACC |
95℃, 11 min |
31 |
95℃, 46 sec |
73℃, 11 min |
|
R:TGAAGCGGTCGGTGAACTG |
63℃, 46 sec |
||||
|
73℃, 2 min 31sec |
ISKpn8: internal region of the ISKpn8 insertion sequence; ISKpn8T: region between ISKpn8 and the truncated blaTEM gene; KPCT: region between the truncated blaTEM gene and the blaKPC-2 gene; F: forward primer; A: reverse primer.
3.7 Dissemination of genetic determinants of resistance
The spread of ESBLs in Enterobacteriaceae is driven by clonal dissemination and mobile genetic elements like plasmids, transposons, and integrons. The blaKPC-2 gene is carried by the Tn3-type transposon Tn4401, which includes tnpA, tnpR, and insertion sequences ISKpn6 and ISKpn7. Tn4401, found in various species and regions, exists in three isoforms (a, b, and c) that differ by 100–200 bp deletions upstream of blaKPC-2, highlighting its high mobility and role in resistance gene spread. Other variants of Tn4401 have been described, mainly involving modifications in the upstream region of blaKPC-2, indicating its instability. In China, a novel genetic environment for blaKPC-2 was identified, featuring an ISKpn6-like sequence downstream of the gene, a newly described ISKpn8 insertion sequence upstream, and a truncated blaTEM gene [37, 38]. For E. coli isolates 1, 2, 3, and 49, which carry blaKPC-2, the presence of Tn4401 elements was investigated using PCR with primers targeting ISKpn7, ISKpn6, tnpA, and tnpR. Only ISKpn6 was amplified, and sequencing confirmed its identity with the previously described Tn4401 transposon. No amplification of tnpA, tnpR, or ISKpn7 was observed. Given this, primers were designed to detect the novel genetic structure described in China, targeting ISKpn8, the region between ISKpn8 and the truncated blaTEM gene, and the region between blaTEM and blaKPC-2, as detailed in Table 7 [37].
Genes upstream of blaKPC-2 could not be identified since no amplification was observed in any of the regions examined. The genetic environment of blaCTX-M-15 was also examined, given its association with mobile elements like the ISEcp1 insertion sequence, which plays a key role in mobilizing blaCTX-M genes. Additional elements, such as IS26 and IS903, also contribute to gene transfer. In E. coli isolates 37 and K. pneumoniae 24, 35, 40, 7, 17, 29, and 43, ISEcp1 was consistently identified upstream of blaCTX-M-15. With the exception of E. coli isolate 37 and K. pneumoniae isolate 24, which exhibited different genetic profiles, IS903 was present. In K. pneumoniae ST15 isolates, both ISEcp1 and IS903 were found flanking blaCTX-M-15, facilitating its mobilization between plasmids and chromosomes. Integron analysis in E. coli isolates 49 and 37, and K. pneumoniae isolates 35, 40, 24, 7, 17, 29, and 43, revealed a class I integron in E. coli isolate 49, containing the dhfrV gene linked to trimethoprim resistance. To investigate plasmid dissemination mechanisms, plasmid incompatibility groups were identified using replicon typing. Five multiplex and three simplex PCR reactions were conducted targeting 18 replication origin groups. E. coli isolates 1, 2, 3, and 49 tested positive for IncFIA, IncFIB, and IncF, while E. coli isolate 37 harbours IncFIA and IncF. Among K. pneumoniae isolates, only isolate 7 carried IncN, with no positive results in others. These findings are consistent with previous research showing that IncFII plasmids are associated with the spread of blaCTX-M-15 in E. coli. At the same time, K. pneumoniae producers of CTX-M β-lactamase are commonly associated with IncFII, IncN, and IncL/M plasmid groups [9].
3.8 Plasmid DNA extraction
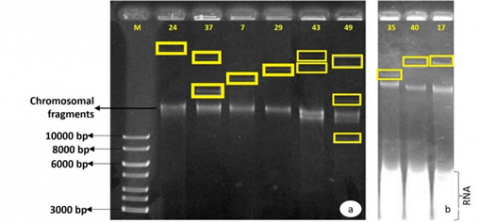
The plasmid DNA extraction technique developed by Kado and Liu [25] was used for E. coli isolates 49 and 37 and K. pneumoniae isolates 24, 35, 40, 7, 17, 29, and 43 to determine the presence of plasmids in bacterial isolates. It was possible to identify plasmids in all isolates studied, with more than one plasmid observed in E. coli isolates 49 and 37 and K. pneumoniae isolate 43 (Figure 5). However, since plasmid DNA molecules can exist in different conformations with varying degrees of supercoiling, these isolates may not contain several plasmids but rather several isoforms of the same plasmid, resulting in the visualization of several bands on the agarose gel. E. coli isolates 49 (carrying a KPC-2 carbapenemase) and 37 (carrying an extended-spectrum β-lactamase CTX-M-15) have two isoforms of the plasmid of the same size, with isolate 49 having a smaller form, approximately 9 kb, which is not found in isolate 37. In K. pneumoniae isolates 35 and 40, which are identical both by the previous typing methods and in terms of the resistance genes they carry (CTX-M-15 and SHV-28), it is possible to identify two plasmids of different sizes, with the size of the plasmid of isolate 40 being similar to that of isolate 17, which carries a CTX-M-15, SHV-1, and TEM-1. The plasmid of isolate 24, which carries the resistance genes CTX-M-15, SHV-12, and TEM-1, is superior to that of the other isolates. This investigation shows the diversity of plasmids found in the various isolates, plasmids that can acquire various resistance genes and serve as a vehicle for their spread [38]. It is possible to observe the chromosomal fragments and RNA.
Figure 5. Visualization of plasmid DNA on 0.7% agarose gel
a: Visualization of plasmid DNA extracted from E. coli and K. pneumoniae; b: Visualization of plasmid DNA extracted from K. pneumoniae; M: GeneRulerTM 1kb DNA ladder molecular mass marker.
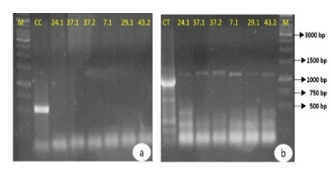
To determine whether the identified resistance genes were plasmid-borne, plasmids were extracted, purified, and analyzed by PCR using DNA recovered from agarose gel. The extracted bands were designated as follows: 37.1 and 37.2 for E. coli 37 (37.1 being the highest molecular mass and 37.2 the lowest), 40.1 for K. pneumoniae 40, 24.1 for K. pneumoniae 24, 35.1 for K. pneumoniae 35, 7.1, 17.1, 29.1, and 43.2 (lowest molecular mass) for K. pneumoniae isolates 7, 17, 29, and 43, and 49.1, 49.2, and 49.3 for E. coli 49 (49.1 being the highest molecular mass and 49.3 the lowest). PCR was conducted to detect blaCTX-M, blaTEM, and blaSHV genes, along with ISEcp1 and IS903 insertion sequences in plasmids 24.1, 37.1, 37.2, 40.1, 35.1, 7.1, 17.1, 29.1, and 43.2. Additionally, blaKPC amplification was performed on plasmids 49.1, 49.2, and 49.3. The results showed that blaTEM was successfully amplified in all tested samples, confirming its plasmid location. However, no amplification was detected for blaCTX-M, blaSHV, or the investigated insertion sequences, suggesting these genes may reside chromosomally rather than on plasmids. Such a transfer may have occurred because ISEcp1 can mobilize blaCTX-M from plasmids to chromosomes. However, the absence of amplification does not definitively exclude plasmid localization, as potential DNA fragmentation during gel extraction may have affected primer binding. Conversely, blaKPC was successfully amplified in plasmids 49.1, 49.2, and 49.3. These findings, particularly the detection of blaTEM in 37.1 and 37.2 and blaKPC in 49.1, 49.2, and 49.3, suggest the presence of different isoforms of the same plasmid rather than multiple distinct plasmids within these isolates. The PCR amplification results for selected samples are shown in Figure 6.
Figure 6. PCR products for the amplification of the 544 bp blaCTX-M (a), 1058 bp blaTEM (b), 886 bp blaSHV (c), and blaKPC (d) genes on 1% agarose gel
CC: positive control for the blaCTX-M gene; CT: positive control for the blaTEM gene; CS: positive control for the blaSHV gene; CK: positive control for the blaKPC gene. M: GeneRulerTM 1kb DNA ladder molecular mass marker.
3.9 Genetic transfer
Genetic transfer techniques, including conjugation and transformation by electroporation, were performed to assess the potential for horizontal transfer of resistance genes among bacterial isolates. Conjugation was attempted in both solid and liquid media using E. coli isolates 1, 3, and 49, which produce KPC-2 carbapenemase; E. coli isolate 37, containing the ESBL-CTX-M-15; and K. pneumoniae isolates 24, 35, 40, 7, 17, 29, and 43, also carrying CTX-M-15. The sodium azide-resistant E. coli J53 strain was used as the recipient. An agar medium supplemented with 1 mg/mL sodium azide was prepared to select transconjugants, and 100 µL of 1 mg/mL cefotaxime was spread onto the surface. However, conjugation was unsuccessful due to issues in transconjugant selection. After 24-hour incubation at 37℃, the observed colonies resembled the donor strain rather than the recipient strain, and unexpected bacterial growth was observed in the negative control, indicating that the sodium azide concentration was insufficient to inhibit donor strain growth, compromising the selection process.
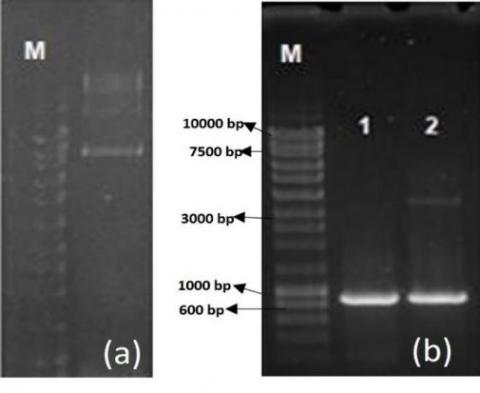
Figure 7. a) Visualization of the plasmid DNA extracted from strain JM109 transformed by electroporation on a 0.7% agarose gel; b) PCR amplification products on 1% agarose gel
1: PCR product for the amplification of the 871 bp blaKPC gene from E. coli isolate 49 (positive control); 2: PCR product for the amplification of the blaKPC gene from transformed strain JM109. M: molecular mass marker NZYDNA ladder III (Nzytech genes & enzymes).
Transformation by electroporation was conducted using E. coli isolates 49 and 37 and K. pneumoniae isolates 24, 40, 7, and 17. Plasmid DNA was extracted using the Kado and Liu [25] method, purified with sodium acetate, and resuspended in sterile water. Competent E. coli JM109 cells were electroporated with the extracted plasmid DNA and incubated at 37℃ for 1 hour before plating on Müller-Hinton agar supplemented with antibiotics to which the donor bacteria were resistant. For E. coli isolate 49, which produces KPC-2 carbapenemase, selection was performed using cefoxitin, ceftazidime, cefotaxime, amoxicillin/clavulanic acid, and imipenem. The ESBL-producing isolates, including E. coli 37 and K. pneumoniae isolates, were selected using cefoxitin, ceftazidime, cefotaxime, and amoxicillin/clavulanic acid. Successful transformation was achieved only for E. coli isolate 49, where the transformed strain exhibited a resistance profile similar to the donor strain, except for gentamicin and ciprofloxacin, for which resistance was not transferred. PCR analysis of DNA extracted from the transformed E. coli JM109 confirmed the presence of the blaKPC gene, and sequencing verified the presence of KPC-2 carbapenemase. Plasmid extraction from the transformed strain revealed a plasmid of approximately 9 kb, further confirmed through PCR as harboring the blaKPC gene (Figure 7).
The plasmid extracted from the transformed strain was analyzed for its origin of replication, identifying IncFIA, IncFIB, and IncF incompatibility groups, also present in the donor strain (E. coli isolate 49). These findings confirm that the blaKPC-2 resistance gene is associated with IncF plasmids, highlighting their role in gene dissemination. Our investigation of two hospitals in Baghdad showed alarming trends in the spread of multidrug-resistant infections. Four genetically identical KPC-2-producing E. coli isolates (1, 2, 3, and 49) were detected at Al-Shaab General Hospital and Al-Yarmouk Teaching Hospital between March 2023 and April 2024. Isolates 1 and 2, recovered from the same patient during separate hospital visits, suggest persistent colonization and nosocomial transmission. Additionally, two clusters of CTX-M-15-producing K. pneumoniae were identified: a 13-isolate group (March–November 2023, represented by isolates 35/40) and a 4-isolate group (April–November 2023, isolates 7/17/29/43). Despite different hospital units, their genetic similarity suggests prolonged circulation and adaptation, underscoring infection control challenges. The extended detection periods (up to eight months) and inter-unit spread highlight the urgent need for enhanced containment measures, including stricter isolation protocols and genomic surveillance. A small sample size, restricted geographic reach, failed conjugation experiments, and potential DNA fragmentation impacting PCR detection are some of the study's limitations. These factors may prevent the study from fully capturing the genetic diversity or resistance mechanisms of Enterobacteriaceae in larger clinical settings.
This study emphasizes the significant prevalence of antibiotic resistance in E. coli and K. pneumoniae isolates from urinary tract infection patients in Baghdad. Resistance rates were highest for ciprofloxacin (92.31%), gentamicin (76.92%), amoxicillin/clavulanic acid (92.31%), ceftazidime (87.18%), and cefotaxime (76.92%), while imipenem showed the highest sensitivity (76.92%). The detection of blaCTX-M-15 in K. pneumoniae ST15 and E. coli ST131, along with blaKPC-2 in E. coli, underscores the role of IncF plasmids in resistance gene dissemination.
The major findings from the study include:
Some specific interventions proposed from the outcome of this study are:
The authors are grateful to the University of Baghdad for their assistance in completing the present research.
[1] MMancuso, G., Midiri, A., Gerace, E., Marra, M., Zummo, S., Biondo, C. (2023). Urinary tract infections: The current scenario and future prospects. Pathogens, 12(4): 623. https://doi.org/10.3390/pathogens12040623
[2] Yang, X., Chen, H., Zheng, Y., Qu, S., Wang, H., Yi, F. (2022). Disease burden and long-term trends of urinary tract infections: A worldwide report. Frontiers in Public Health, 10: 888205. https://doi.org/10.3389/fpubh.2022.888205
[3] Morocco, E., Ferzandi, T. (2022). Urinary tract infections and asymptomatic bacteriuria. In Handbook of Gynecology, pp. 1-9. https://doi.org/10.1007/978-3-319-17002-2_95-1
[4] Alkan, S., Balkan, I.I., Surme, S., Bayramlar, O.F., et al. (2024). Urinary tract infections in older adults: Associated factors for extended-spectrum beta-lactamase production. Frontiers in Microbiology, 15: 1384392. https://doi.org/10.3389/fmicb.2024.1384392
[5] Younus, N.K. (2024). Phenotypic and genotypic characterization of multidrug-resistant Escherichia coli and Klebsiella pneumoniae isolated from women with urinary tract infections in Mosul City. Iraqi Journal of Science, 65(1): 24-35. https://doi.org/10.24996/ijs.2024.65.1.3
[6] Raheem, T.F., Hasan Ali, S.A. (2022). Prevalence and multi-drug resistance patterns of uropathogenic E. coli isolated from women patients in Kirkuk City, Iraq. Iranian Journal of Medical Microbiology, 16(6): 609-614. https://doi.org/10.30699/ijmm.16.6.609
[7] Husna, A., Rahman, M.M., Badruzzaman, A.T.M., Sikder, M.H., Islam, M.R., Rahman, M.T., Alam, J., Ashour, H.M. (2023). Extended-Spectrum β-Lactamases (ESBL): Challenges and Opportunities. Biomedicines, 11(11): 2937. https://doi.org/10.3390/biomedicines11112937
[8] Polse, R.F., Yousif, S.Y., Assafi, M.S. (2016). Prevalence and antimicrobial susceptibility patterns of uropathogenic E. coli among people in Zakho, Iraq. International Journal of Research in Medical Sciences, 4(4): 1219-1223. https://doi.org/10.18203/2320-6012.ijrms20160813
[9] Bevan, E.R., Jones, A.M., Hawkey, P.M. (2017). Global epidemiology of CTX-M β-lactamases: Temporal and geographical shifts in genotype. Journal of Antimicrobial Chemotherapy, 72(8): 2145-2155. https://doi.org/10.1093/jac/dkx146
[10] Al-Dahmoshi, H.O.M., Al-Khafaji, N.S.K., Al-Allak, M.H.O. (2020). Antibiotic resistance among Iraqi local E. coli isolates. E. Coli Infections - Importance of Early Diagnosis and Efficient Treatment. https://doi.org/10.5772/intechopen.92107
[11] Kadeřábková, N., Mahmood, A.J.S., Mavridou, D.A.I. (2024). Antibiotic susceptibility testing using minimum inhibitory concentration (MIC) assays. NPJ Antimicrobials and Resistance, 2: 37. https://doi.org/10.1038/s44259-024-00051-6
[12] Khan, Z.A., Siddiqui, M.F., Park, S. (2019). Current and emerging methods of antibiotic susceptibility testing. Diagnostics, 9(2): 49. https://doi.org/10.3390/diagnostics9020049
[13] Algarni, S., Gudeta, D.D., Han, J., Nayak, R., Foley, S.L. Genotypic analyses of IncHI2 plasmids from enteric bacteria. Scientific Reports, 14: 9802. https://doi.org/10.1038/s41598-024-59870-2
[14] Sharma, A., Lee, S., Park, Y.S. (2020). Molecular typing tools for identifying and characterizing lactic acid bacteria: A review. Food Science and Biotechnology, 29: 1301-1318. https://doi.org/10.1007/s10068-020-00802-x
[15] Zheng, X., Xu, D., Yan, J., Qian, M., Wang, P., Zaeim, D., Han, J., Qu, D. (2024). Mobile genetic elements facilitate the transmission of antibiotic resistance genes in multidrug-resistant Enterobacteriaceae from duck farms. Food Science and Human Wellness, 13: 729-735. https://doi.org/10.26599/FSHW.2022.9250062
[16] Bello-López, J.M., Cabrero-Martínez, O.A., Ibáñez-Cervantes, G., Hernández-Cortez, C., Pelcastre-Rodríguez, L.I., Gonzalez-Avila, L.U., Castro-Escarpulli, G. (2019). Horizontal gene transfer and its association with antibiotic resistance in the genus Aeromonas spp. Microorganisms, 7(9): 363. https://doi.org/10.3390/microorganisms7090363
[17] Michaelis, C., Grohmann, E. (2023). Horizontal gene transfer of antibiotic resistance genes in biofilms. Antibiotics, 12(2): 328. https://doi.org/10.3390/antibiotics12020328
[18] Wellinghausen, N., Kochem, A., Disqué, C., Mühl, H., Gebert, S., Winter, J., Matten, J., Sakka, S.G. (2009). Diagnosis of bacteremia in whole-blood samples by use of a commercial universal 16S rRNA gene-based PCR and sequence analysis. Journal of Clinical Microbiology, 47(9): 2759-2765. https://doi.org/10.1128/JCM.00567-09
[19] Chela, H.K., Vasudevan, A., Rojas-Moreno, C., Naqvi, S.H. (2019). Approach to positive blood cultures in the hospitalized patient: A review. Missouri Medicine, 116(4): 313-317.
[20] Gajic, I., Kabic, J., Kekic, D., Jovicevic, M., Milenkovic, M., Mitic Culafic, D., Trudic, A., Ranin, L., Opavski, N. (2022). Antimicrobial susceptibility testing: A comprehensive review of currently used methods. Antibiotics, 11(4): 427. https://doi.org/10.3390/antibiotics11040427
[21] Guinea, J. (2020). Updated EUCAST clinical breakpoints against Aspergillus: Implications for the clinical microbiology laboratory. Journal of Fungi, 6(4): 343. https://doi.org/10.3390/jof6040343
[22] Kockum, I., Huang, J., Stridh, P. (2023). Overview of genotyping technologies and methods. Current Protocols, 3(4): e727. https://doi.org/10.1002/cpz1.727
[23] Jacob, M.S., Kulie, P., Benedict, C., Ordoobadi, A.J., Sikka, N., Steinmetz, E., McCarthy, M.L. (2018). Use of a midstream clean catch mobile application did not lower urine contamination rates in an ED. The American Journal of Emergency Medicine, 36(1): 61-65. https://doi.org/10.1016/j.ajem.2017.07.016
[24] Ghaly, T.M., Chow, L., Asher, A.J., Waldron, L.S., Gillings, M.R. (2017). Evolution of class 1 integrons: Mobilization and dispersal via food-borne bacteria. PloS One, 12(6): e0179169. https://doi.org/10.1371/journal.pone.0179169
[25] Kado, C.I., Liu, S.T. (1981). Rapid procedure for detection and isolation of large and small plasmids. Journal of Bacteriology, 145(3): 1365-1373. https://doi.org/10.1128/jb.145.3.1365-1373.1981
[26] Matsumura, Y., Peirano, G., Pitout, J. (2018). Complete genome sequence of Escherichia coli J53, an azide-resistant laboratory strain used for conjugation experiments. Genome Announcements, 6(21): e00433-18. https://doi.org/10.1128/genomeA.00433-18
[27] Jomehzadeh, N., Ahmadi, K., Bahmanshiri, M.A. (2022). Investigation of plasmid-mediated quinolone resistance genes among clinical isolates of Klebsiella pneumoniae in southwest Iran. Journal of Clinical Laboratory Analysis, 36(7): e24342. https://doi.org/10.1002/jcla.24342
[28] Lee, M.M.Y., Kuan, N.L., Li, Z.Y., Yeh, K.S. (2024). Occurrence and characteristics of extended-spectrum-β-lactamase- and pAmpC-producing Klebsiella pneumoniae isolated from companion animals with urinary tract infections. PLoS ONE, 19(1): e0296709. https://doi.org/10.1371/journal.pone.0296709
[29] Oraby, H.A.S., Aboul-Maaty, N.A.F., Al-Sharawi, H.A., Osman, N.M. (2022). Horizontal transfer of antibiotic resistance genes into microflora and blood cells in rats fed on GM-diet. Bulletin of the National Research Centre, 46(1): 268. https://doi.org/10.1186/s42269-022-00956-0
[30] Pérez-Palacios, P., Gual-de-Torrella, A., Portillo-Calderón, I., Recacha-Villamor, E., Franco-Álvarez de Luna, F., Lopez-Cerero, L., Pascual, A. (2023). Interhospital spread of blaVIM-1- and blaCTX-M-15-Producing K. pneumoniae ST15 on an IncR Plasmid in Southern Spain. Antibiotics, 12(12): 1727. https://doi.org/10.3390/antibiotics12121727
[31] Dan, T., Liu, W., Sun, Z., Lv, Q., Xu, H., Song, Y., Zhang, H. (2014). A novel multi-locus sequence typing (MLST) protocol for Leuconostoc lactis isolates from traditional dairy products in China and Mongolia. BMC Microbiology, 14: 150. https://doi.org/10.1186/1471-2180-14-150
[32] Forde, B.M., Roberts, L.W., Phan, M.D., Peters, K.M., et al. (2019). Population dynamics of an Escherichia coli ST131 lineage during recurrent urinary tract infection. Nature Communications, 10(1): 3643. https://doi.org/10.1038/s41467-019-11571-5
[33] Gondal, A.J., Saleem, S., Jahan, S., Choudhry, N., Yasmin, N. (2020). Novel carbapenem-resistant klebsiella pneumoniae ST147 coharboring bla NDM-1, bla OXA-48 and extended-spectrum β-lactamases from Pakistan. Infection and Drug Resistance, 2020: 2105-2115. https://doi.org/10.2147/IDR.S251532
[34] Brown, N.G., Shanker, S., Prasad, B.V., Palzkill, T. (2009). Structural and biochemical evidence that a TEM-1 β-lactamase N170G active site mutant acts via substrate-assisted catalysis. Journal of Biological Chemistry, 284(48): 33703-33712. https://doi.org/10.1074/jbc.M109.053819
[35] Higgins, O., Chueiri, A., O'Connor, L., Lahiff, S., et al. (2023). Portable differential detection of CTX-M ESBL gene variants, bla CTX-M-1 and bla CTX-M-15, from Escherichia coli isolates and animal fecal samples using loop-primer endonuclease cleavage loop-mediated isothermal amplification. Microbiology Spectrum, 11(1): e03316-22. https://doi.org/10.1128/spectrum.03316-22
[36] Hong, J.H., Clancy, C.J., Cheng, S., Shields, R.K., Chen, L., Doi, Y., Zhao, Y., Perlin, D.S., Kreiswirth, B.N., Nguyen, M.H. (2013). Characterization of porin expression in Klebsiella pneumoniae carbapenemase (KPC)-producing K. pneumoniae identifies isolates most susceptible to the combination of colistin and carbapenems. Antimicrobial Agents and Chemotherapy, 57(5): 2147-2153. https://doi.org/10.1128/AAC.02411-12
[37] Chen, L., Chavda, K.D., Melano, R.G., Jacobs, M.R., Levi, M.H., Bonomo, R.A., Kreiswirth, B.N. (2013). Complete sequence of a blaKPC-2-harboring IncFIIK1 plasmid from a Klebsiella pneumoniae sequence type 258 strain. Antimicrobial Agents and Chemotherapy, 57(3): 1542-1545. https://doi.org/10.1128/AAC.02332-12
[38] Rozwandowicz, M., Brouwer, M.S.M., Fischer, J., Wagenaar, J.A., Gonzalez-Zorn, B., Guerra, B., Mevius, D.J., Hordijk, J. (2018). Plasmids carrying antimicrobial resistance genes in Enterobacteriaceae. Journal of Antimicrobial Chemotherapy, 73(5): 1121-1137. https://doi.org/10.1093/jac/dkx488