Severe Overexploitation of the Mud Spiny Lobster (panulirus polyphagus) Revealed by Length-Based Assessment in Gresik, East Java, Indonesia

Arief Setyanto*![]() | Ledhyane Ika Harlyan
| Ledhyane Ika Harlyan![]() | Viga Ballacka Irawan | Putri Farmita Maryam | Hanif Rabbani Rama Putra | Romeo Abimanyu Kusuma Wardhana | Andik Isdianto
| Viga Ballacka Irawan | Putri Farmita Maryam | Hanif Rabbani Rama Putra | Romeo Abimanyu Kusuma Wardhana | Andik Isdianto![]() | Agus Tumulyadi
| Agus Tumulyadi![]() | Nico Rahman Caesar
| Nico Rahman Caesar![]() | Uun Yanuhar
| Uun Yanuhar![]()
© 2026 The authors. This article is published by IIETA and is licensed under the CC BY 4.0 license (http://creativecommons.org/licenses/by/4.0/).
OPEN ACCESS
A critical lack of local biological and population data continues to undermine sustainable management of lobster fisheries in Indonesia. This study aims to assess the population dynamics of lobsters caught by the fishers through the identification of the lobster species, describe their life history traits, and evaluate their exploitation status. The data collected included morphological characteristics, carapace length (CL), body weight, Gonad Maturity Level (GML), East Java fisheries production data, and sea surface temperature (SST). Length-frequency data were analyzed to estimate growth parameters (using ELEFAN I) and to assess exploitation status using the length-based spawning potential ratio (LB-SPR) approach. The study identified three lobster species: Panulirus polyphagus, Panulirus ornatus, and Panulirus versicolor. Species composition was dominated by P. polyphagus (95%) with a total of 467 individuals (237 males and 230 females). The length-weight relationship of P. polyphagus exhibited a negative allometric growth pattern. The estimated length at first maturity (Lm) was 86.49 mm, while the length at first capture (Lc) was 69.5 mm, indicating that the average size of caught P. polyphagus individuals did not reach gonadal maturity (Lc < Lm). The asymptotic length was 92.18 mm with a growth coefficient of 1.5/year. The LB-SPR value for P. polyphagus was 3%, which falls under the overexploited category. Therefore, a reduction in fishing effort for lobster is recommended in the northern waters of Gresik.
lobster, Gresik, length-based spawning potential ratio, northern coast of Java, Pantura
Indonesia has a variety of fisheries resources, one of which is the lobster (Panulirus spp.) or spiny lobster. Lobsters are crustaceans with hard exoskeletons that inhabit coral and rocky reefs, as well as sandy or muddy areas. Coral and rocky reefs not only provide habitat but also serve as feeding grounds [1, 2]. Lobster species belong to the family Palinuridae, consisting of 49 species, with 11 species found in the Indo-Pacific region, 6 of which are found in Indonesian and East Java waters [3-6]. Six species of spiny lobster with different morphologies include the ornamental lobster (Panulirus ornatus), Pakistan lobster (P. polyphagus), rock lobster (P. penicillatus), sand lobster (P. homarus), batik lobster (P. longipes), and bamboo lobster (P. versicolor). Lobsters have high economic value, which shows that among nine commodities with the highest economic value, lobster has the highest value for five consecutive years compared to other crustacean species such as crabs. East Java is one of the provinces in Indonesia known for lobster production. The lobster produced comes from various bio-ecoregions in East Java, one of which is Gresik Regency, located on the northern coast of the region [4]. The lobsters harvested in the Northern Coast of East Java and the Madura Strait are landed by local small-scale fishermen. The lobster catch is considered bycatch of swimming crab gears such as gillnets and traps [7, 8]. This differs from the southern coast of East Java, where lobsters are a primary target species [8]. Fish Landing Bases (PPI) are the places where fishermen land their bycatch. Gresik Regency has five PPIs spread across the northern region, consisting of PPI Pangkah Wetan, PPI Tirta Buana, PPI Kalinggapuri, PPI Campurejo, and PPI Cabean. Although there are five PPIs, only two PPIs handle lobster landings, namely PPI Pangkah Wetan and PPI Campurejo. The landed lobsters come from fishing activities by local fishermen, but there is no proper record-keeping of lobster production data, making it difficult to know the exact number and species of lobsters caught during each fishing operation. The lack of clear record-keeping results in a shortage of information on the number and species of lobsters that have been exploited, leading to minimal information regarding the utilization status of lobsters’ resources in Gresik Waters. This issue could result in over-exploitation of lobster fisheries due to the lack of information on utilization status, causing continuous fishing without applying sustainable practices. This lack prevents quantitative population assessments, thus hindering the development of scientific management benchmarks (such as MSY, FMSY). This study will fill this gap by applying the data-limited assessment method (LB-SPR).
In small-scale and bycatch-dominated fisheries, limited data availability often restricts the application of conventional stock assessment approaches [9, 10]. The Length-Based spawning potential ratio (LB-SPR) is a stock assessment framework based on population reproductive theory that estimates the remaining spawning potential of an exploited population relative to its unfished state using body length data, key life-history parameters, and fishing mortality inferred from catch size structure. Due to its strong biological basis, low data requirements, and broad application in data-limited fisheries, the LB-SPR framework is particularly suitable for assessing exploitation status in small-scale fisheries. Accordingly, this study applies the LB-SPR framework to evaluate the exploitation status of spiny lobster (Panulirus spp.) in Gresik Waters [11-13]. Therefore, this study aims to: (1) identify the dominant spiny lobster species and describe their population size structure in Gresik Waters; (2) estimate key life-history parameters derived from length-based data; and (3) assess the current exploitation status of spiny lobster resources using the LB-SPR framework. We hypothesize that the dominant species, particularly Panulirus polyphagus, may be experiencing a high level of exploitation in Gresik Waters.
2.1 The study area
The research was conducted over a period of three months (January – April 2023) at PPI Campurejo and PPI Pangkah Wetan, Gresik Regency (Figure 1). Data collection was carried out through observation, interviews, and documentation.

Figure 1. Map of study area: PPI Ujung Pangkah and Campurejo, Gresik, East Java
2.2 Procedure
2.2.1 Data collection
Sampling was conducted at PPI Campurejo and PPI Pangkah Wetan, Gresik Regency, from January to April 2023. Lobster sampling was performed 2–3 times per week following the landing schedule of small-scale fishing vessels, with each sampling event covering the entire landing period (approximately 3–5 hours). All lobsters landed during each sampling event were measured whenever possible. When landings were large, random subsampling was applied to obtain representative length-frequency data. Carapace length (CL, mm) was measured using a digital caliper, and sex was determined based on external morphological characteristics. A total of 467 individual lobsters were recorded during the study period. This number represents the cumulative sample collected across all sampling events and does not reflect the total population size. Monthly sample sizes varied in accordance with fishing intensity. Lobsters were captured as bycatch of small-scale fisheries targeting swimming crabs, primarily using gillnets and traps. Fishing effort was characterized based on the number of fishing trips and landing events observed, with an average of 8–15 vessels landing catches per sampling day, and each vessel conducting a single-day fishing trip.
2.3 Data analysis
2.3.1 Descriptive analysis
Descriptive analysis is the summarization and explanation of the characteristics of the data, such as the description of lobster species. Descriptive analysis is conducted by examining the morphology of lobsters found in Gresik Waters. Morphological analysis includes physical characteristics, body shape, and the color of the lobsters caught. The species composition was then measured using the percentage formula.
2.3.2 Relationship between carapace length and lobster weight
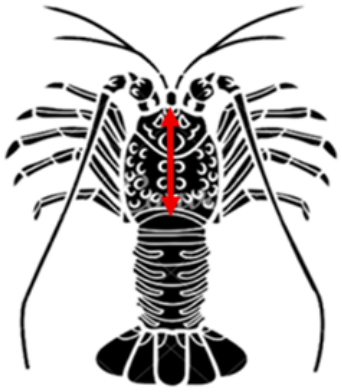
CL was measured as the distance from the posterior margin of the eye socket to the midpoint of the posterior margin of the carapace along the dorsal midline. Measurements were recorded to the nearest 0.01 mm using a digital caliper following standard morphometric procedures for spiny lobsters. The length-frequency data were subsequently used to estimate key life-history parameters and the spawning potential ratio (SPR) of the lobster population [14-17]. A schematic illustration of the CL measurement is provided in Figure 2 to ensure measurement consistency.
Figure 2. Schematic illustration of carapace length (CL) measurement (the red double-headed arrow) in spiny lobster (Panulirus spp.)
CL is measured as the linear distance from the posterior margin of the eye socket to the midpoint of the posterior margin of the carapace along the dorsal midline.
The estimation of the relationship between the CL and the weight of lobster species is performed using the cubic model, Eq. (1) [9].
$W=a . C L^b$ (1)
where,
W = Weight (g)
CL = Carapace length (mm/cm)
a = Constant
b = Allometric constant
The value of b is then tested using a T-test with the following Eq. (2).
$\begin{aligned} { thit } & =\frac{b-3}{s e} ; \\ t_{t a b} & =t_{\frac{\alpha}{2}, n-2}\end{aligned}$ (2)
where,
b = The regression coefficient (slope) obtained from the length–weight relationship analysis, commonly expressed in the Eq. (1).
se = The standard error of the coefficient b, representing the uncertainty or variability of the estimated slope value.
thit = t calculated = The calculated t-statistic used to test whether the estimated parameter b is significantly different from the theoretical value.
tₜₐb(t-tab) = The critical value of the t-distribution obtained from the t-table, used as a comparison value for the calculated t-statistic in hypothesis testing.
α (alpha) = The significance level of the statistical test (e.g., 0.05 or 0.01). It represents the probability of rejecting the null hypothesis when it is actually true.
α/2 = The half of the significance level, used in a two-tailed test because the rejection region is divided into two tails of the distribution.
n = The sample size or number of observations used in the analysis.
Hypothesis:
H0: b = 3, meaning the relationship between length and weight is isometric.
H1: b ≠ 3, meaning the relationship between length and weight is allometric.
If thit < ttab, then accept H0 and reject H1.
If thit > ttab, then reject H0 and accept H1.
2.3.3 Sex
The sex ratio is estimated using Eq. (3) [10].
$N K=\frac{\sum J}{\sum B}$ (3)
where,
NK = Sex ratio
$\sum J$ = Number of male lobsters
$\sum B$ = Number of female lobsters
The results are then tested using the chi-square test (X²) with a 95% confidence interval (p = 0.05) to test for uniformity [11]. The calculation uses Eq. (4).
$X^2=\Sigma \frac{\left(O_i-E_i^2\right)}{E_i}$ (4)
where,
O = Observed frequency
E = Expected frequency
2.3.4 Length at first capture (Lc)
The estimation of the first capture size is based on the distribution of length frequency, which is analyzed using a normal distribution equation. The mathematical model uses Eq. (5) as follows [12]:
$\mathrm{F}_{\mathrm{c}}=(\mathrm{ndL} / \mathrm{s} \sqrt{2} \pi)^* \mathrm{e}\left\{-\left(\mathrm{L}^{\prime \prime}-\mathrm{L}\right)^2 / 2 \mathrm{~s}^2\right\}$ (5)
where,
Fc = frequency of fish in length classes
n = number of samples in the sampling
dL = length of class interval
s = standard deviation
π = constant 3.14
L’’ = midpoint value of the length class
L = average length of a fish cohort
The estimation of the average and standard deviation of fish length is performed by converting the Eq. (5) into a linear form as Eq. (6) follows:
$\Delta \ln F_c(z)=a-b \times(L+d L / 2)$ (6)
where,
∆lnFc(z) = The change in the natural logarithm of corrected catch frequency for the z-th length class. This value is used in catch curve analysis to estimate mortality rates.
L + dL/2 = Corrected midpoint of the length class, used to better represent the actual position of the length class in the regression analysis.
a, b = intercept and slope coefficient
The mean length and standard deviation of each specific age group are estimated using Eq. (7).
$\mathrm{Lc}=\mathrm{a} / \mathrm{b}$ and $\mathrm{s}^2=-\mathrm{dL} / \mathrm{b}$ (7)
2.3.5 Length at first mature (Lm)
Gonadal maturity was classified using a five-stage Gonadal Maturity Stage (GMS) system, as presented in the Results section (GMS I–V). The stages were defined as follows: GMS I (Immature), gonads small and undeveloped; GMS II (Developing), gonads enlarged but translucent; GMS III (Mature), gonads clearly developed with increased coloration; GMS IV (Ripe), gonads fully developed and intensely colored; and GMS V (Spent), gonads flaccid or partially resorbed after spawning. Gonadal maturity was determined through macroscopic examination following standard references for spiny lobster (Panulirus spp.) [15].
For analytical purposes, individuals categorized as GMS III–V were considered mature and were used in the estimation of length at first maturity (Lm).
The estimation of Lm is calculated using Microsoft Excel with the following Eq. (8) [10].
$L n=\left(\frac{1-p}{p}\right)=r(L m-r L)$ (8)
where,
p = Proportion of fish with mature gonads
r = Slope (regression result of the proportion of mature gonads and fish length)
L = Mean length class value of the fish (mm)
Lm = Average length at first gonad maturity (mm)
Ln = Data regression
The estimation of P or the probability of the estimation is calculated using the following Eq. (9).
$P=\frac{1}{\left(1+e^{-r(L-L m)}\right)}$ (9)
The value of L95 can be calculated using Eq. (10) from Prince et al. [13] as follows:
$L 95=1.1 \times L 50$ (10)
2.3.6 Length-based spawning potential ratio
The estimation of natural mortality (M) is calculated using the empirical Eq. (11) as follows [13]:
$Log \,\,M=(-0,0066)-0,279 \,\, Log \,\, {L}{\infty}+0,6453 \,\,Log \,K+0,4634 \,\,Log \,\,T$ (11)
where,
M = Natural mortality
L∞ = Asymptotic length (mm)
K = Growth coefficient
T = Average temperature during the study, which was 29 ℃.
The asymptotic length (L∞) and growth coefficient (K) are obtained using FISAT II with the ELEFAN I method. The value of t0 can be determined using the empirical Eq. (12) [14].
$Log \left(-t_0\right)=-0.3922-0.2752 Log L_{\infty}-1.038 \log (K)$ (12)
where,
L∞ = Asymptotic length
K = Growth coefficient
The growth rate can be estimated using the Von Bertalanffy model as follows [12]:
$\mathrm{Lt}=\mathrm{L} \infty\left(1-e^{-k\left(t-t_0\right)}\right)$ (13)
where,
Lt = lobster length at age t (mm)
L∞ = asymptotic length (mm)
t0 = theoretical age when the lobster's length is 0
K = growth coefficient
The average sea surface temperature (SST) was obtained by downloading Aqua MODIS data from the Ocean Color website, which was then processed using Seadas and ArcMap, resulting in a temperature (T) value of 29 ℃.
The estimation of the LB-SPR value can be performed using Eq. (14) from Hordyk et al. [15]:
$S P R=\frac{S S B R_{ {fished }}}{S S B R_{{unfished }}}$ (14)
where,
SSBRfished = Spawning stock biomass per recruit under fishing
SSBRunfished = Spawning stock biomass per recruit in the unfished state
The LB-SPR value can be analyzed using the LB-SPR method through the Barefoot Ecologist website by selecting the LB-SPR menu.
3.1 Lobster species identification
The species found in the northern waters of Gresik consist of three species, namely P. polyphagus, P. ornatus, and P. versicolor (Tables 1 and 2). The morphological characteristics of each species differ. The key differences can be observed in the antennular plate and abdominal somites of each species.
Table 1. Various names of lobster species caught in Gresik Waters
|
Name |
Mud Spiny Lobster |
Ornate Spiny Lobster |
Painted Spiny Lobster |
|
Scientific Name |
Panulirus polyphagus |
Panulirus ornatus |
Panulirus versicolor |
|
Indonesia Name |
Lobster pakistan |
Lobster mutiara |
Lobster bambu |
|
Local Name |
Udang jarak |
Udang ketangan |
Udang karang |
Table 2. Lobster species of the Gresik waters of East Java
|
|
Lobster Pakistan (Panulirus polyphagus) |
|
|
Lobster Bambu (Panulirus versicolor) |
|
|
Lobster Mutiara (Panulirus ornatus) |
3.2 Species composition
The species caught were dominated by P. polyphagus with 467 individuals (95%), consisting of 237 males and 230 females, while P. ornatus accounted for 16 individuals (3%), with 8 males and 8 females. P. versicolor consisted of 8 individuals (2%), with 3 males and 5 females (Figure 3). The dominance of P. polyphagus is the reason why this study uses P. polyphagus as the subject; thus, several subsequent analyses include sex ratio analysis, length-weight relationship, length at first maturity, Lc, asymptotic length, growth coefficient, natural mortality, and LB-SPR using the species P. polyphagus.
3.3 Sex ratio
The sex ratio between males and females resulted in a comparison ratio of 1:1, with a chi-square test showing X² calculated = 0.00035 and X² table = 3.84146 (X² calculated < X² table), indicating that there is no significant difference between the proportions of male and female lobsters (Table 3). Therefore, for subsequent analysis, female lobsters were used as the subject. The analysis conducted includes the relationship between length and weight, Lc, Lm, asymptotic length, growth coefficient, natural mortality, and LB-SPR.
3.4 The length frequency distribution of lobster carapace
P. polyphagus observed during the study had a length class range between 47 and 89 mm. The most dominant length class for P. polyphagus was 65 mm, with 95 individuals caught. The fewest individuals were found in the 83 mm length class. The age group distribution from the length frequency of P. polyphagus shows one age group in each month (Figure 4).
The CL frequency of P. ornatus shows the highest capture in the 67.1 mm length class with 6 individuals. The 79.1 mm and 103.1 mm length classes each had 1 individual. The age groups in the graph indicate that there is only 1 age group in January (Figure 5). In February and March, no age group was found because only 1 individual was captured in each of those months. In April, no individuals were found, so the age group graph for that month is not displayed.
The P. versicolor observed during the study had a length class range between 62.2 and 72.2 mm. The dominant average catch occurred at the length classes of 62.2 mm and 72.2 mm, with 2 individuals caught at each length. One individual was caught at a length of 67.2 mm. The age group of P. versicolor showed the presence of one age group in January, while no age group was found in February, March, and April, as P. versicolor was not found during those months (Figure 6).
Table 3. Male and female sex ratio
|
Species |
Number |
Total |
Sex Ratio |
Percentage |
Expected |
X2cal |
X2table |
|||
|
Male |
Female |
Male |
Female |
Male |
Female |
|||||
|
P. polyphagus |
237 |
230 |
467 |
1 |
1 |
51% |
49% |
233.5 |
0.00035 |
3.84146 |
|
|
|
|
a |
b |
Figure 3. Composition of lobster species in the catch (a) Total lobster; (b) Female lobster

Figure 4. Monthly length–frequency distribution of sampled individuals of P. polyphagus from January to April 2023

Figure 5. Length frequency distribution of P. ornatus for samples collected in January 2023

Figure 6. Length frequency distribution P. versicolor for samples collected in January 2023
3.5 Relationship between length and weight
The relationship between length and weight for P. polyphagus is expressed by the equation W = 0.0578 × CL1.9961, where Fsig ≤ 0.001, a = 0.0578, and b = 1.9961. The result of the t-test shows t = 14.34, df = 465, p < 0.001, indicating that H0 is rejected and H1 is accepted. This means that the growth pattern of P. polyphagus is negatively allometric. The graph showing the relationship between CL and weight can be seen in Figure 7.
3.6 Length at first capture and length at first maturity
The calculation of length at first maturity (Lm) in this study uses female lobster CL data along with Gonad Maturity Stage (GMS) data, while the calculation of Lc uses female lobster CL data. The Lm and Lc graphs can be seen in Figure 8.

Figure 7. Relationship between carapace length (CL) and weight of P. polyphagus
The results of the Lm and Lc graphs show that the Lc value is 69.5 mm and the Lm value is 86.49 mm. The value of Lc < Lm indicates that the average lobster caught has not yet reached gonadal maturity or has not yet reproduced.
|
|
|
|
(a) |
(b) |
Figure 8. Graph of Lm and Lc for P. polyphagus (a) length at first maturity (Lm); (b) length at first capture (Lc)
3.7 Length-based spawning potential ratio
The LB-SPR analysis requires life history values, including asymptotic length, growth coefficient, natural mortality, as well as Lm50 and Lm95 values. The life history results can be seen in the following table (Table 4). The difference between Lm95 and L∞ is relatively small (≈3 mm), indicating that the estimated maturity size is close to the asymptotic length.
Table 4. Life history
|
Parameter |
Estimate (unit) |
|
M |
3.61/Year |
|
K |
1.5/ Year |
|
L∞ |
92.18 (mm) |
|
Lm50 |
86.49 (mm) |
|
Lm95 |
95.14 (mm) |
The life history results were then input into the Barefoot Ecologist website, yielding an LB-SPR value of 3%, which falls into the over-exploited category. A graph of the LB-SPR value can be seen in Figure 9.

Figure 9. Length-based spawning potential ratio (LB-SPR) value graph
Life-history parameters derived for P. polyphagus included an asymptotic length (L∞) of 92.18 mm, a growth coefficient (K) of 1.5 year⁻¹, and a natural mortality rate (M) of 3.61 year⁻¹. The estimated reproductive parameters were Lm₅₀ = 86.49 mm and Lm₉₅ = 90 mm (Table 4). Using these parameters, the LB-SPR analysis yielded an SPR value of 3% (Figure 9). This value indicates that the current spawning biomass of P. polyphagus in the study area is approximately 3% of the unfished spawning potential, reflecting a severely depleted reproductive stock under the existing fishing regime.
The species composition of lobsters caught in Gresik waters differs from that in other regions of East Java, such as Southern Java, Madura Strait, and Madura archipelago [4, 6, 7]. The species composition in the Southern Java region is of 6 species: P. homarus, P. penicillatus, P. longipes, P. versicolor, P. ornatus, and P. polyphagus, respectively [5, 6, 16]. In North Java, 4 species are found: P. polyphagus, P. ornatus, P. versicolor, and P. homarus [4, 7]. The 4 species were also found in the Madura Strait region but with different species composition, i.e., P. homarus, P. ornatus, P. versicolor, and P. polyphagus, respectively [7]. The species of P. ornatus, P. versicolor, P. polyphagus, P. homarus, P. longipes, and P. penicillatus are found in the Madura Archipelago [4-8]. The differences in species composition of Indonesia archipelago found in each region can be attributed to variations in the evolutionary geological history among marine regions [17, 18].
Negative allometric growth patterns were also observed in studies in Karimunjawa with the species P. versicolor, Situbondo with P. ornatus, and Sebatik with P. polyphagus [7, 19, 20]. Similar growth patterns can occur due to similarities in environmental characteristics that support the availability of food and suitable habitats [21, 22]. The negative allometric growth pattern observed in Panulirus polyphagus (b < 3) reflects a shift in energy allocation strategy, where individuals invest proportionally less energy in somatic mass relative to linear growth. Rather than being solely attributed to environmental variation, this pattern likely represents an adaptive response to ecological stress and intense fishing pressure, in which energy is prioritized toward maintenance and reproduction rather than body mass accumulation. Persistent size-selective fishing may further reinforce this pattern through fishing-induced selection, favoring phenotypes that mature earlier or allocate less energy to somatic growth, thereby increasing survival under high exploitation regimes.
The Lm value for P. polyphagus in Gresik waters is Lm = 86.49 mm. This value is much higher compared to the Lm value in the Strait of Johor, which is Lm = 65.9 mm [23]. Another study in Sebatik waters, North Kalimantan, found a Lm value of 90.74 mm. The differences in Lm values in each study may be influenced by the fishing pressure in the respective waters [24]. The asymptotic length (L∞) in Gresik waters is smaller (L∞ = 92.18 mm) compared to an L∞ value of 134.4 mm in Sebatik waters, North Kalimantan [25]. This difference is believed to be due to variations in environmental characteristics [26]. The differences can also be caused by the period of sample collection, sample size, food abundance, fishing pressures and environmental conditions in the study area [22, 24].
The exploitation status of P. polyphagus is over-exploited in Gresik waters, which was also observed in the study conducted in Sebatik waters, North Kalimantan, with an LB-SPR value of 19%. Over-exploitation in the utilization status can indicate overfishing of the species. In addition to indicating overfishing, an over-exploited utilization status may also indicate that the immature gonads and small size of the catches are dominated. Management of P. polyphagus utilization in Gresik waters needs to be carried out to prevent extinction.
Research on the life history and population dynamics of lobsters in North Gresik waters can be compared with similar studies from various regions in Indonesia. One study conducted in Gunungkidul waters, Yogyakarta, analyzed the population structure and biological aspects of lobsters, particularly Panulirus penicillatus and Panulirus homarus. The study showed a negative allometric growth pattern for both lobster species. Additionally, the SPR for P. homarus was recorded at 17%, which is categorized as over-exploited, while P. penicillatus had an SPR of 30%, which falls under the moderate category [27-29]. Study highlights the importance of more sustainable management to prevent over-exploitation of lobster populations in the region.
Another study was conducted in the Madura Strait, Situbondo, East Java, aiming to determine the species composition, length frequency distribution, growth patterns, and sex ratio of lobsters [7]. This research provides additional insights into the population dynamics of lobsters in East Java waters. The information gathered can complement data from Gresik waters to understand the distribution patterns and population structure of lobsters along East Java's coastal areas as a whole. This indicates that regional studies can provide a more comprehensive understanding. Thus, this study is relevant as a comparison for analyzing the biological aspects [30, 31] lobsters’ population in Gresik.
A study in Pelabuhanratu waters, West Java, also provides insights into lobster population dynamics, specifically of the species of P. penicillatus and P. homarus [32, 33]. This study examined growth, recruitment patterns, mortality, and exploitation rates of sand lobsters. The results showed that the exploitation rate of the species had exceeded the optimal level, indicating overfishing activities. This phenomenon is consistent with findings from other regions, indicating high fishing pressure on lobster populations. By comparing the results of your study in Gresik with research in Palabuhanratu, similar patterns in exploitation and their impacts on lobster population sustainability are obvious.
Additionally, a study conducted in Kebumen assessed the utilization status of lobster species and found that continuous fishing activities had impacted the sustainability of the lobster population in the area [30, 31, 34]. This study noted that uncontrolled fishing activities could significantly reduce the number of adult lobsters. This aligns with the importance of implementing ecosystem-based management policies to maintain the balance of lobster populations [35]. Comparing the research findings with data from Kebumen will provide a broader context regarding the implications of lobster exploitation in various coastal regions of Indonesia.
The condition where the (Lc = 69.5 mm CL) is smaller than the (Lm₅₀ = 86.49 mm CL) indicates that Panulirus polyphagus in Gresik waters is predominantly harvested before reproduction, resulting in truncated size structure and reduced spawning stock biomass. This ecological impact is quantitatively confirmed by the LB-SPR of 3%, indicating that the stock retains only 3% of its unfished reproductive potential and is therefore in a critically overexploited state. Such an extremely low SPR implies that fishing mortality strongly exceeds natural mortality (F/M ≫ 1), identifying fishing pressure as the dominant driver of population decline. Compared with other LB-SPR studies on spiny lobsters and similar species, which typically report SPR values of 17–30% under moderate to high exploitation, the value observed in this study is among the lowest reported. SPR < 20% is generally considered the threshold for overfishing, while 3% means that the reproductive potential has been basically lost, and the population may face the risk of collapse if no urgent measures are taken. The concurrent occurrence of negative allometric growth (b < 3) and relatively high Lm should not be attributed solely to environmental variation but likely reflects adaptive energy allocation strategies and fishing-induced selection, where individuals invest less in somatic growth or delay maturation under intense size-selective harvesting. Together, these findings demonstrate that unsustainable fishing pressure has profoundly altered the life-history traits and reproductive capacity of P. polyphagus in Gresik waters, underscoring the urgent need for management measures to reduce fishing pressure and protect mature individuals.
Research on the life history and population dynamics of lobsters in Gresik waters can be compared with similar studies from other regions of Indonesia, providing a more comprehensive understanding of the distribution and population structure of lobsters along East Java's coastal areas. The findings from other regions, such as Pelabuhan Ratu and Kebumen, show similar patterns of exploitation, underscoring the importance of sustainable management practices to maintain lobster population sustainability across Indonesia’s coastal regions.
The species composition of lobsters in Gresik waters differs from that of other regions in East Java, such as Southern Java, the Madura Strait, and the Madura Archipelago, reflecting the variations in lobster species diversity across different marine regions. Several studies have observed negative allometric growth patterns in species such as P. versicolor, P. ornatus, and P. polyphagus, likely influenced by similar environmental characteristics that support the availability of food and suitable habitats.
The Lm value for P. polyphagus in Gresik waters is significantly higher compared to other regions, possibly influenced by varying fishing pressures. Differences in L∞ values and exploitation status indicate environmental factors that affect the development of this species. In Gresik waters, P. polyphagus is over-exploited, which may indicate overfishing and the predominance of immature individuals, highlighting the urgent need for sustainable management to prevent the species from facing depletion.
[1] Amin, M., Harlyan, L.I., Achmad, K., Diantari, R. (2022). Profiling the natural settlement habitat of spiny lobster, Panulirus spp. to determine potential diets and rearing conditions in a lobster hatchery. Biodiversitas: Journal of Biological Diversity, 23(6): 2893-2898. https://doi.org/10.13057/biodiv/d230615
[2] Konecny, C.A., Cote, D., Broome, J., Nicolas, J.M., Regular, P.M., Cook, A.M., Hatefi, F. (2024). Influences of environmental and individual-level covariates on movement behaviour in American lobster Homarus americanus. Marine Ecology Progress Series, 729: 151-166. https://doi.org/10.3354/meps14519
[3] George, R.W. (1998). Tectonic plate movements and the evolution of Jasus and Panulirus spiny lobsters (Palinuridae). Marine and Freshwater Research, 48(8): 1121-1130. https://doi.org/10.1071/MF97202
[4] Setyanto, A., Rachman, N.A., Yulianto, E.S. (2019). Distribution and composition of lobster species caught in Java Sea of East Java, Indonesia. Jurnal Perikanan Universitas Gadjah Mada, 20(2): 49-55.
[5] Setyanto, A., Soemarno, Wiadnya, D.G.R., Nugroho, C. (2019). Biodiversity of lobster (Panulirus) from eastern Indian Ocean of Indonesia waters. IOP Conference Series: Materials Science and Engineering, 546(2): 022024. https://doi.org/10.1088/1757-899X/546/2/022024
[6] Setyanto, A., Soemarno, D.G.R., Prayogo, C., West, R.J., Tsamenyi, M., Halimah, S., Rahman, N.A. (2020). Spiny lobsters species composition between North and South Sea of East Java, Indonesia. Ecology, Environment & Conservation, pp. S49-S53.
[7] Setyanto, A., Sambah, A.B., Widhiastika, D., Soemarno, Wiadnya, D.G.R., Prayogo, C. (2021). Population structure and biological aspects of lobster (Panulirus spp.) of the Madura Strait landed in Situbondo of East Java, Indonesia. IOP Conference Series: Earth and Environmental Science, 919(1): 012015. https://doi.org/10.1088/1755-1315/919/1/012015
[8] Setyanto, A., Wiadnya, D.G.R., Prayogo, C., Kusuma, Z., West, R.J., Tsamenyi, M. (2023). Fishing methods and fishing season of the tropical lobster fisheries of Southern Java, Indonesia. Biodiversitas: Journal of Biological Diversity, 24(2): 776-783. https://doi.org/10.13057/biodiv/d240213
[9] Sparre, P. (1998). Introduction to Tropical Fish Stock Assessment: Part 1. Manual. FAO Fisheries Technical Paper.
[10] King, M. (2007). Fisheries Biology, Assessment and Management. John Wiley & Sons.
[11] Zar, J.H. (1999). Biostatistical Analysis. Pearson Education India.
[12] Sparre, P. (1998). Introduction to tropical fish stock assessment. Part 1. Manual. FAO Fisheries Technical Paper, 306: 1-407. https://doi.org/10.2331/suisan.78.681
[13] Prince, J.D., Wilcox, C., Hall, N. (2023). How to estimate life history ratios to simplify data-poor fisheries assessment. ICES Journal of Marine Science, 80(10): 2619-2629.
[14] Pauly, D. (1983). Some Simple Methods for the Assessment of Tropical Fish Stocks (Vol. 234). Food & Agriculture Org.
[15] Hordyk, A., Ono, K., Valencia, S., Loneragan, N., Prince, J. (2015). A novel length-based empirical estimation method of spawning potential ratio (SPR): And tests of its performance, for small-scale, data-poor fisheries. ICES Journal of Marine Science, 72(1): 217-231. https://doi.org/10.1093/icesjms/fsu004
[16] Setyanto, A., Soemarno, Wiadnya, D.G.R., Nugroho, C. (2019). Biodiversity of lobster larvae (Panulirus spp.) from the Indonesian Eastern Indian Ocean. IOP Conference Series: Earth and Environmental Science, 370: 012046. https://doi.org/10.1088/1755-1315/370/1/012046
[17] Ferrari, D.S., Floeter, S.R., Leprieur, F., Quimbayo, J.P. (2023). A trait-based approach to marine island biogeography. Journal of Biogeography, 50(3): 528-538. https://doi.org/10.1111/jbi.14549
[18] Komai, T., Yang, C.H., Chan, T.Y. (2020). Deep-sea shrimps of the genus Glyphocrangon A. Milne-Edwards, 1881 (Decapoda: Caridea: Glyphocrangonidae) collected by the SJADES 2018 expedition off Java, Indonesia, with description of one new species. Raffles Bulletin of Zoology, 68: 636-653. https://doi.org/10.26107/RBZ-2020-0079
[19] Ernawati, T., Priatna, A., Satria, F. (2019). Biological reference points of painted spiny lobster Panulirus versicolor (Latreille, 1804) in Karimun Jawa waters, Indonesia. Indonesian Fisheries Research Journal, 25(2): 91-101. https://doi.org/10.15578/ifrj.25.2.2019.91-101
[20] Suman, A., Chodrijah, U., Kang, B., Zhang, C.I. (2021). Stock assessment and management implications of three lobster species in Gunungkidul waters, Indonesia. Ocean & Coastal Management, 211: 105780. https://doi.org/10.1016/j.ocecoaman.2021.105780
[21] Kawirian, R.R., Affandi, R., Mashar, A., Effendi, I. (2023). Food preferences of early juvenile scalloped spiny lobster in Ekas Bay, Lombok, Indonesia. Indonesian Aquaculture Journal, 18(1): 61-69. https://doi.org/10.15578/iaj.18.1.2023.61-69
[22] Radhakrishnan, E.V., Phillips, B.F., S, L.P., Padua, S. (2019). Ecology and global distribution pattern of lobsters. Lobsters: Biology, Fisheries and Aquaculture, pp. 151-176. https://doi.org/10.1007/978-981-32-9094-5_5
[23] Waiho, K., Fazhan, H., Shu-Chien, A.C., Abualreesh, M.H., et al. (2021). Size distribution, length-weight relationship, and size at morphometric maturity of the mud spiny lobster Panulirus polyphagus (Herbst, 1793) in the Johor Strait. Frontiers in Marine Science, 8: 766038. https://doi.org/10.3389/fmars.2021.766038
[24] Chodrijah, U., Priatna, A., Nugroho, D. (2018). Length size distribution and population parameters of mud lobster (Panulirus polyphagus Herbst, 1793) in Sebatik Waters, North Kalimantan (WPPNRI-716). Jurnal Penelitian Perikanan Indonesia, 24(1): 11-23. https://doi.org/10.15578/jppi.1.1.2018.11-23
[25] Tirtadanu, T., Chodrijah, U., Wagiyo, K. (2021). Reference point and exploitation status of mud spiny lobster (Panulirus polyphagus Herbst, 1793) in Sebatik waters, Indonesia. Indonesian Fisheries Research Journal, 27(1): 27-36. https://doi.org/10.15578/ifrj.27.1.2021.27-36
[26] Yusuf, Z.A., Abdulkarim, M. (2015). Fundamentals of tropical fish stock assessment.
[27] Lestari, P.A., Zairion, Z., Mashar, A., Widianto, T. (2024). Lobster fisheries management strategy by the social-ecological system approach: The case of Gunungkidul Coastal Waters, Special Region of Yogyakarta Province. BIO Web of Conferences, 112: 11005. https://doi.org/10.1051/bioconf/202411211005
[28] Kembaren, D.D., Pane, A.R.P., Wagiyo, K., Mardlijah, S. (2024). Assessing risk status for some commercial fisheries in Gunungkidul waters, Indonesia: An ecosystem-based fisheries approach. Egyptian Journal of Aquatic Research, 50(1): 141-147. https://doi.org/10.1016/j.ejar.2024.01.001
[29] Suman, A., Chodrijah, U., Zhang, C.I. (2022). Multi-species assessment and management implications of lobster fisheries in Gunungkidul waters, Indonesia. Egyptian Journal of Aquatic Research, 48(1): 91-98. https://doi.org/10.1016/j.ejar.2021.10.006
[30] Irwani, Sabdono, A., Wijayanti, D.P. (2019). Growth, mortality and exploitation rate of spiny lobster (Panulirus homarus) from Kebumen and Cilacap coastal. IOP Conference Series: Earth and Environmental Science, 246(1): 012081. https://doi.org/10.1088/1755-1315/246/1/012081
[31] Widianti, E.A., Nurani, T.W., Sondita, M.F.A., Purwangka, F., Wahyuningrum, P.I. (2021). Catch composition of lobsters (Panulirus spp) at Karangduwur Landing site in Kebumen Regency, Central Java. Albacore, 5(2): 121-132. https://doi.org/10.29244/core.5.2.121-132
[32] Boer, M., Wahyudin, R.A., Wardiatno, Y., Farajallah, A., Hakim, A.A. (2021). Population dynamics of pronghorn spiny lobster (Panulirus penicillatus) in Palabuhanratu Bay, Sukabumi, West Java. https://doi.org/10.29244/jpsl.11.2.204-214
[33] Zairion, Z., Islamiati, N., Wardiatno, Y., Mashar, A., Wahyudin, R.A., Hakim, A.A. (2017). Population dynamics of scalloped spiny lobster (Panulirus homarus Linnaeus, 1758) in Palabuhanratu Waters, West Java. Jurnal Penelitian Perikanan Indonesia, 23(3): 215-226.
[34] Wahyuningrum, P.I., Widianti, E.A., Kurniawati, V.R., Citra Bangun, T.N., Nurani, T.W. (2022). Type and catch size of lobsters (Panulirusspp) landed at Fisheries Landing Centres in Kebumen Regency. Albacore, 6(3): 323-332. https://doi.org/10.29244/core.6.3.323-332
[35] Widhiastika, D., Taurusman, A.A., Wahju, R.I. (2024). Management status of the lobster (Panulirus spp.) fisheries based in Prigi Bay, Trenggalek, East Java: A human dimension of ecosystem approach. Egyptian Journal of Aquatic Biology and Fisheries, 28(4): 765-781. https://doi.org/10.21608/ejabf.2024.369523