Population Structure and Regeneration Dynamics of the Endangered Ebony (Diospyros celebica Bakh.) Across Distinct Ecological Gradients in South Sulawesi, Indonesia

Samuel A. Paembonan*![]() | Merryana Kiding Allo
| Merryana Kiding Allo![]() | Syamsuddin Millang
| Syamsuddin Millang![]() | Budi Arty
| Budi Arty![]() | Ahmad Rifqi Makkasau
| Ahmad Rifqi Makkasau![]() | Resti Ura’
| Resti Ura’![]() | Nardy Noerman Najib
| Nardy Noerman Najib![]() | Dicky
| Dicky![]()
© 2025 The authors. This article is published by IIETA and is licensed under the CC BY 4.0 license (http://creativecommons.org/licenses/by/4.0/).
OPEN ACCESS
Diospyros celebica Bakh. is an endangered Sulawesi endemic increasingly confined to fragmented, selectively logged forests, yet its population demography across natural habitats remains poorly known. This study quantified population structure, regeneration dynamics, and species associations of D. celebica along edaphic and disturbance gradients in three natural stands in South Sulawesi (Maros, Barru, Sidrap). Vegetation was sampled using belt transects with nested quadrats across four life stages (seedlings, saplings, poles, trees) and analysed using phytosociological indices, important value index (IVI), diversity metrics, soil properties, and chi-square association tests. In Maros, ebony showed strong dominance in tree and pole layers and substantial juvenile representation, forming a size class distribution closely approximating a reverse J-shaped curve (power regression: y = 711.98x⁻²·⁶⁶⁸), indicative of continuous regeneration under moderately acidic, organic-rich soils. Barru exhibited extremely dense seedlings but sharp declines at sapling and tree stages (y = 2312.7x⁻²·¹¹⁴), coinciding with more acidic, low-organic soils and intense competition from shade-tolerant palms. In Sidrap, ebony remained the main canopy species but was strongly underrepresented in juvenile strata (y = 605.8x⁻¹·¹⁵⁸) dominated by other tree taxa, signalling an aging, demographically eroding population. Site-specific association patterns, particularly negative relationships with dense understorey palms, highlight biotic filters on early regeneration. These findings support differentiated conservation strategies tailored to each stand type to secure the long-term persistence of D. celebica in South Sulawesi’s tropical forests.
conservation management, endangered species, natural forest, population structure, regeneration dynamics, South Sulawesi, species association, Wallacea region
Tropical forests are among the most biodiverse ecosystems on Earth, yet they continue to face unprecedented pressure from land-use change, overharvesting, and habitat degradation [1-3]. In Indonesia, these pressures have severely impacted populations of island-endemic, high-value timber species [4]. Among such species, the black ebony, Diospyros celebica Bakh. (Ebenaceae), endemic to Sulawesi, produces highly prized, dark-coloured timber used in furniture, handicrafts, and decorative applications [5-7]. Its economic value has driven intensive and often unsustainable logging [8], while natural regeneration has not kept pace with extraction [9].
The black ebony, Diospyros celebica Bakh., is an endemic tree species of Sulawesi that produces high-quality, dark-colored timber widely used for furniture, handicrafts, and decorative purposes [9-11]. Its high economic value has driven intensive and often unsustainable logging [12], while natural regeneration has not kept pace with extraction, raising serious concerns about the long-term viability of wild populations [7, 13]. D. celebica is currently listed as endangered and is increasingly restricted to fragmented and degraded habitats [14]. Recent studies have addressed aspects of its potential distribution, habitat suitability, and autecology, but most have focused on limited geographic areas, such as North and Central Sulawesi seed stands [13, 15], and have not fully examined population structure and community context across multiple natural habitats.
South Sulawesi, particularly the districts of Maros, Barru, and Sidenreng Rappang (Sidrap), encompasses diverse ecological conditions within the Wallacea region—a global biodiversity hotspot and centre of endemism [10, 11]. This region harbours distinct combinations of climate, topography, soils, and disturbance regimes that have shaped the distribution and regeneration dynamics of tropical forest communities [12]. The three sites selected for this study represent contrasting natural habitats of D. celebica, capturing gradients from relatively intact stands to more disturbed or stressed ecosystems. Understanding how ebony populations respond across these ecological gradients is essential for developing evidence-based conservation strategies in a region where deforestation and selective logging continue to fragment natural forests [13].
By integrating population structure, important value index (IVI), soil properties, and species association analyses, the study aims to provide an ecological diagnosis of the current status of D. celebica and its co-occurring species, and to derive implications for sustainable management and conservation.
2.1 Study area
The study was conducted in three natural ebony habitats located in Maros, Sidrap, and Barru districts, South Sulawesi Province, Indonesia (Figure 1). The sites were purposively selected to represent contrasting combinations of climate, topography, soils, and disturbance history, thereby capturing a range of ecological conditions for Diospyros celebica.
All sites lie within the tropical humid zone, from lowland to lower montane elevations, with mean annual rainfall of approximately 1,500–2,500 mm and mean temperatures of 26–29℃ (Table 1).
Table 1. Environmental description at the 3 research locations
|
Parameter |
Maros |
Sidrap |
Barru |
|
Coordinate point |
4°58′26.22″ S, 119°45′59.23″ E |
3°44′33.49″ S, 120°03′29.52″ E |
4°26′01.34″ S, 119°38′19.00″ E |
|
Rainfall (mm year⁻¹) |
2,000–2,500 |
1,500–2,500 |
1,500–2,500 |
|
Elevation (m a.s.l.) |
550–604 |
96–220 |
90–250 |
|
Topography |
Undulating to steep topography |
gentle to hilly |
Undulating |
|
Slope gradient (%) |
30–45 |
20–40 |
20–40 |
|
Temperature (℃) |
27–28 |
26–29 |
27–29 |
|
Relative humidity (%) |
75–85 |
70–90 |
74–90 |

Figure 1. Map of research location in South Sulawesi, Indonesia
2.2 Sampling design and data collection
Vegetation was sampled using continuous belt transects with nested quadrats at each site (Figure 2). Three transects of 600 m length were established at each site, along which main plots of 20 × 20 m (400 m²) were established at regular continuous sample plots to record all tree-layer individuals (diameter at breast height (DBH) ≥ 20 cm), resulting in a total of 30 main plots per site (3 transects × 10 plots per transect = 4,000 m² of tree-layer sampling per site). Within each tree plot, nested subplots were laid out as follows: 10 × 10 m (100 m²) for poles (DBH 10–20 cm), 5 × 5 m (25 m²) for saplings (DBH < 10 cm and height > 1.5 m), and 2 × 2 m (4 m²) for seedlings (height < 1.5 m). In each plot or subplot, all woody individuals meeting the size criteria were identified to species, counted, and measured for DBH (poles and trees) and total height; clear bole height was additionally recorded for trees and poles. For saplings and seedlings, abundance and height were recorded.

Figure 2. Form of sampling plot on the observation line
Soil samples from the upper mineral layer (0–30 cm) were collected at 30 fixed sampling points per site (one per main plot), with each sampling point consisting of a composite sample of three soil cores taken within the vegetation plot. This ensured spatial correspondence between soil and vegetation data.
2.3 Environmental conditions and soil analyses
Environmental characteristics and soil properties at the three research sites were analyzed to determine the environmental conditions that support the growth of the surrounding vegetation communities. The environmental variables analyzed in this study were those related to climate, including rainfall, temperature, and relative humidity. Other factors included topography, slope, and elevation (Table 1).
Table 2. Physical and chemical properties of soil at the research location
|
Chemical Properties |
Units |
Barru |
Sidrap |
Maros |
|
pH (H₂O) |
– |
4.79 ± 0.06 |
5.95 ± 0.11 |
5.40 ± 0.21 |
|
C-organic |
% |
1.30 ± 0.05 |
1.67 ± 0.14 |
2.21 ± 0.34 |
|
N-total |
% |
0.14 ± 0.01 |
0.16 ± 0.01 |
0.12 ± 0.01 |
|
P-available |
ppm |
10.22 ± 0.73 |
10.43 ± 0.80 |
19.25 ± 3.11 |
|
Exchangeable K |
ppm |
8.28 ± 0.42 |
10.12 ± 0.70 |
9.76 ± 0.79 |
|
CEC |
cmol(+)·kg-1 |
0.21 ± 0.01 |
0.21 ± 0.02 |
0.22 ± 0.01 |
|
Physical Properties |
Units |
Barru |
Sidrap |
Maros |
|
Soil Texture |
– |
Clay loam |
Sandy loam |
Sandy loam |
|
Sand |
% |
47.63 ± 1.02 |
45.10 ± 3.67 |
63.33 ± 2.95 |
|
Silt |
% |
32.53 ± 0.81 |
33.10 ± 2.58 |
24.50 ± 2.35 |
|
Clay |
% |
19.84 ± 0.71 |
21.90 ± 2.04 |
12.00 ± 1.13 |
|
Porosity |
% |
7.33 ± 0.19 |
7.17 ± 0.32 |
6.96 ± 0.60 |
Soil samples were analysed for pH (H₂O), organic carbon, total nitrogen, available phosphorus, exchangeable potassium, cation exchange capacity (CEC), texture (sand, silt, clay), and porosity using standard laboratory procedures. These variables were used to characterise edaphic gradients among sites (Table 2).
2.4 Data analysis
Statistical analysis of soil differences among sites
One-way ANOVA was performed on each soil variable (pH, organic carbon, total nitrogen, available phosphorus, exchangeable potassium, CEC, sand/silt/clay fractions, and porosity) to test for significant differences among the three sites. Levene's test was used to check homogeneity of variance; where variance homogeneity was violated, Kruskal–Wallis non-parametric tests were performed instead. Where significant differences were detected (p < 0.05), post-hoc pairwise comparisons were conducted using Tukey's HSD test (parametric) or Dunn's test (non-parametric) to identify which site pairs differed significantly. All statistical tests were performed using R version 4.0 (R Core Team, 2023).
Phytosociological parameters and IVI
For each species and growth stratum (trees, poles, saplings, seedlings), basic phytosociological parameters were calculated as:
$Density(\mathrm{D})=\frac{N_i}{A}$
where, $N_i$ is the total number of individuals of species i recorded in the sampling plots, and A is the total sampled area (ha).
$Relative density \left(R D_i\right)=\frac{D_i}{\sum D} \times 100$
where, $R D_i$ is the relative density of species $i(\%), D_i$ is the density of species $i$, and $\sum \mathrm{D}$ is the sum of densities of all species within the same growth stage.
$Frequency \left(F_i\right)=\frac{\text { number of plots where species } i \text { occurs }}{\text { total number of plots }}$
$Relative frequency\left(R F_i\right)=\frac{F_i}{\sum F} \times 100$
where, $R F_i$ is the relative frequency of species $i(\%), F_i$ is the frequency of species i, and $\sum \mathrm{F}$ is the sum of frequencies of all species.
For poles and trees, basal area BA was computed as:
$B A_i=\sum \frac{\pi D B H^2}{4}$
where, BAi is the total basal area of species i (m²), π is a constant (3.1416), and DBH is the diameter at breast height (1.3 m above ground) of each individual tree or pole (cm).
And dominance and relative dominance were:
$Dominance \left(D o_i\right)=\frac{B A_i}{A}$,
where Doi is the dominance of species i (m² ha⁻¹), $B A_i$ is the total basal area of species i, and A is the total sampled area (ha).
$Relative\ dominance(R D o)=\frac{D o_i}{\sum D o} \times 100$
where, RDo is the relative dominance of species i (%), $D o_i$ is the dominance of species i, and ∑Do is the sum of dominance values of all species.
The IVI was then derived as:
$IVI_{\text {trees, poles, saplings }}=R D+R F+R D o$
$\mathrm{IVI}_{\text {seedlings }}=R D+R F$
Diversity, richness, and evenness
Species diversity was quantified using the Shannon–Wiener index:
$H^{\prime}=-\sum_{i=1}^S p_i \ln p_i$
where, $S$ is the total number of species, and $p_i=n_i / N$ is the proportion of individuals of species $i\left(n_i\right)$ relative to the total number of individuals $(N)$.
Species evenness was calculated using Pielou’s evenness index:
$J^{\prime}=\frac{H^{\prime}}{\ln S}$
where, H' is the Shannon–Wiener diversity index, S is the total number of species, and ln denotes the natural logarithm.
Species richness was estimated using Margalef’s index:
$D_{M g}=\frac{S-1}{\ln N}$
where, DMg is Margalef’s richness index, N is the total number of individuals recorded.
Population structure of Diospyros celebica
The population structure of D. celebica was described by plotting the density of individuals per hectare across the four growth stages (seedlings, saplings, poles, trees) for each site to assess whether size‑class distributions approximated a reverse J‑shaped curve indicative of continuous regeneration.
Species association analysis
Species associations between D. celebica and other taxa were analysed using $2 \times 2$ contingency tables at the subplot level. For each pair of species, a contingency table was constructed with cell frequencies $a, b, c$, and $d$ representing the number of sampling units where: both species are present (a); only species A is present (b); only species B is present (c); and both are absent (d); with $N=a+b+c+d$ as the total number of sampling units.
The significance of the association was tested with the chi‑square statistic (with Yates’ continuity correction):
$\chi^2=\frac{\left(|a d-b c|-\frac{N}{2}\right)^2 N}{(a+b)(c+d)(a+c)(b+d)}$
where, a, b, c, and d represent the observed frequencies in each cell of the 2 × 2 contingency table, and N is the total number of observations. The term |ad − bc| denotes the absolute difference of cross-products, while N/2 is the Yates’ continuity correction, applied to reduce overestimation of statistical significance when sample sizes are small.
The calculated $\chi^2$ value was compared with the critical value at 1 degree of freedom and $\alpha=0.05$ to determine whether the association was statistically significant.
The strength and direction of association were quantified using the Ochiai index:
$I O=\frac{a}{\sqrt{(a+b)(a+c)}}$
where, a is the number of joint occurrences of two species, b is the number of occurrences of the first species without the second, and c is the number of occurrences of the second species without the first. The Ochiai index ranges from 0 to 1, with higher values indicating a stronger association or similarity between the two species.
Values of IO were classified into categories from very low to very high association strength following research [16-19].
3.1 Environmental and soil characteristics
The three study sites are all located in the tropical humid zone of South Sulawesi but differ in climate and topographic setting. All three sites are located in tropical lowland to lower montane zones but differ in climatic and edaphic conditions (Tables 1 and 2). Maros is situated at elevations around 550 m above sea level with steep, dissected hill slopes, whereas Barru spans a slightly broader altitudinal range (approximately 90–250 m) and includes more gently sloping to hilly terrain. Mean annual rainfall is generally medium across all sites (about 1,500–2,500 mm year⁻¹), and mean temperatures range from 26 to 29℃, conditions that are broadly favourable for the development of evergreen and semi-evergreen tropical forest. The combination of high rainfall and steep slopes in Maros promotes rapid drainage and can increase susceptibility to erosion, while the more varied relief in Barru creates a mosaic of microhabitats that may differentially affect tree establishment and growth.
Soil analyses revealed clear edaphic contrasts among the three sites. All soils were acidic to slightly acidic, but Barru exhibited the lowest pH (4.79 ± 0.06), significantly lower than both Sidrap (5.95 ± 0.11) and Maros (5.40 ± 0.21) (p < 0.01). Organic carbon content followed a distinct gradient, being highest in Maros (2.21 ± 0.34%), intermediate in Sidrap (1.67 ± 0.14%), and lowest in Barru (1.30 ± 0.05%); C-organic did not differ significantly between Barru and Sidrap (p = 0.101) but was markedly higher in Maros compared with both sites (p < 0.001). This pattern was broadly reflected in soil quality indicators, as Maros also exhibited relatively higher CEC, although differences in CEC among sites were not statistically significant (p > 0.05). Total nitrogen varied only slightly, with Sidrap (0.16 ± 0.01%) showing significantly higher values than Maros (0.12 ± 0.01%; p = 0.022), while Barru (0.14 ± 0.01%) did not differ from either site. In contrast, the C/N ratio was substantially higher in Maros (19.25 ± 3.11) than in Barru (8.28 ± 0.42) and Sidrap (10.43 ± 0.80) (p < 0.001), indicating slower organic matter turnover. Available phosphorus, exchangeable potassium, and soil moisture content showed no significant differences among locations. Texturally, Barru soils were dominated by clay loam, whereas Sidrap and Maros were characterized by sandy loam, with Maros having a significantly higher sand fraction (63.3 ± 3.0%) than Barru and Sidrap (p < 0.001), which in turn exhibited higher silt and clay contents (p < 0.05). Porosity values indicated generally suboptimal structural conditions across all sites, with Barru showing the poorest porosity, while Sidrap and Maros displayed moderately improved but still constrained soil structure. Collectively, these patterns suggest that Maros combines moderately acidic conditions with higher organic carbon, wider C/N ratios, and coarser texture, potentially enhancing nutrient retention and rooting conditions, whereas Barru’s more acidic, organic-poor, and structurally less favorable soils may impose stronger limitations on nutrient availability and early plant establishment, with Sidrap representing an intermediate edaphic condition.
3.2 Population structure of Diospyros celebica
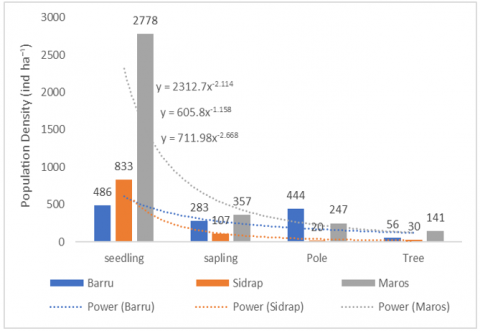
Figure 3 focuses specifically on the population structure of Diospyros celebica and shows how the density of ebony individuals changes from seedlings through saplings and poles to trees at each site. The population structure of D. celebica varied markedly among the three sites. In Maros, ebony showed high densities across all growth stages, with numerous seedlings and saplings and substantial numbers of poles and trees, forming a size‑class distribution close to a reverse J-shaped curve [20].
Figure 3. Population structure of D. celebica
In Barru, ebony seedlings were very abundant, but densities declined sharply in the sapling and tree stages, while the pole stage remained well represented, indicating a bottleneck between early regeneration and the transition to mature trees.
This suggests that, when seedlings do establish successfully, they may grow relatively well into intermediate size classes, yet the transition from poles to mature trees is less efficient, possibly due to selective logging, competition, or site limitations.
In Sidrap, ebony was abundant only in the tree layer, with relatively low densities in all earlier stages, implying poor recruitment and a risk of future decline in canopy dominance. D. celebica shows low densities in all juvenile stages and is mainly represented by adult trees, which indicates a strongly aging population with limited ongoing recruitment. This skewed structure, dominated by older size classes, is a warning sign that, without targeted management interventions, ebony may gradually lose its dominant position in the canopy as current adult trees senesce and die without sufficient replacement.
3.3 Community structure of the natural ebony ecosystem
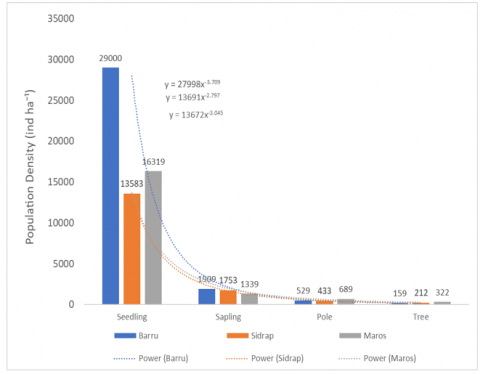
Figure 4 illustrates the density of all woody species recorded in the natural ebony ecosystem across the four growth stages (seedlings, saplings, poles, and trees) at the three study sites. Overall, the number of individuals is highest in the seedling layer and declines progressively towards the tree layer at all sites, which is consistent with the general expectation of a reverse J-shaped structure in regenerating tropical forests.
Figure 4. Community structure of the natural D. celebica ecosystem
Despite this common tendency, the magnitude of decline between stages differs among sites, indicating distinct regeneration dynamics of the community as a whole. At Barru, the extremely high density of seedlings followed by a sharp reduction at the sapling and tree stages suggests intense thinning and high mortality during early development, even though the pole layer remains relatively well represented. In Sidrap, a more even distribution among saplings, poles, and trees, combined with lower overall densities, points to a community under stronger disturbance or with generally lower regeneration capacity. In Maros, the smoother decline from seedlings to trees, with comparatively high densities maintained in all stages, indicates a more continuous and balanced recruitment process, and thus a community structure closer to a stable, self-replacing forest.
Taken together, the tendencies depicted in Figures 3 and 4 show that, while the broader woody community at all sites retains a generally declining pattern from seedlings to trees, the specific trajectory of D. celebica differs markedly among locations. Maros represents a relatively well-functioning ebony population embedded in a regenerating community, Barru exhibits a bottleneck between intermediate and adult stages, and Sidrap already shows signs of demographic decline, with regeneration insufficient to sustain future canopy dominance.
3.4 Vegetation structure of ebony communities (IVI)
At the Barru site, the tree layer was co-dominated by Intsia bijuga and Diospyros celebica, which showed similarly high IVI values of 61.72 and 59.50, respectively (Table 3). This dual dominance indicates that ebony plays a major ecological role in the canopy, even though it does not form a monospecific stand. In the pole layer, D. celebica was clearly the most dominant species, with an IVI of 141.93 that far exceeded all other species, reflecting a strong recruitment of individuals into the intermediate growth stage. In contrast, the seedling layer was dominated by understory palms and herbs such as Arenga pinnata and Calamus sp., while D. celebica showed only a very low IVI (2.68), suggesting serious constraints at the earliest regeneration stage.
Table 3. Dominant species based on the important value index (IVI) in the natural D. celebica community at Barru
|
Growth Stratum |
Species |
IVI |
|
Tree |
Intsia bijuga |
61.72 |
|
Diospyros celebica |
59.50 |
|
|
Arenga pinnata |
36.71 |
|
|
Artocarpus sericicarpus |
23.20 |
|
|
Koordersiodendron pinnatum |
10.63 |
|
|
Pole |
Diospyros celebica |
141.93 |
|
Knema pubiflora |
22.03 |
|
|
Knema celebica |
18.28 |
|
|
Garcinia sp. |
9.49 |
|
|
Filicium decipiens |
7.79 |
|
|
Sapling |
Diospyros celebica |
25.60 |
|
Lasianthus firmus |
23.14 |
|
|
Garcinia sp. |
13.53 |
|
|
Knema pubiflora |
11.06 |
|
|
Mangifera foetida |
10.02 |
|
|
Seedling |
Arenga pinnata |
35.53 |
|
Calamus sp. |
17.76 |
|
|
Donax cannaeformis |
17.61 |
|
|
Licuala rumphii |
14.80 |
|
|
Diospyros celebica |
2.68 |
In Sidrap, D. celebica was the key canopy species, with the highest IVI (73.31) in the tree layer, whereas several co-occurring species, such as Knema tomentella, Payena acuminata, and Endiandra rubescens, had only moderate IVI values (about 17–20) (Table 4). This pattern indicates that the current overstorey is still strongly structured by ebony, but the community remains relatively diverse. However, in the pole and sapling layers, dominance shifted towards other species, particularly Payena acuminata and Shorea koordesii, while ebony appeared only with low to moderate IVI. In the seedling layer, regeneration was largely dominated by non-ebony species, including E. rubescens and Amorphophallus konjac, which suggests a potential future shift in stand composition away from ebony if current trends continue.
At Maros, D. celebica exhibited very strong dominance in both the tree and pole layers, with IVI values of 119.07 and 129.80, respectively, much higher than those of other species such as Arenga pinnata and Mangifera quadrifida (Table 5). This clearly shows that ebony acts as a true keystone species in structuring the main forest canopy at this site. In the sapling and seedling layers, ebony still maintained relatively high IVI values but faced increasing competition from species such as Cinnamomum iners, Areca catechu, and Palaquium obovatum. Overall, the vertical distribution of IVI at Maros suggests a more stable population structure, in which strong dominance in the upper strata is still supported by meaningful regeneration in the lower layers.
Table 4. Dominant species based on the important value index (IVI) in the natural D. celebica community at Sidrap
|
Growth Stratum |
Species / Local Name |
IVI |
|
Tree |
Diospyros celebica |
73.31 |
|
Knema tomentella |
19.91 |
|
|
Payena acuminata |
19.20 |
|
|
Endiandra rubescens |
18.57 |
|
|
Diospyros lotus |
17.25 |
|
|
Pole |
Payena acuminata |
42.54 |
|
Shorea koordesii |
23.80 |
|
|
Knema tomentella |
18.82 |
|
|
Endiandra rubescens |
18.50 |
|
|
Diospyros lotus |
16.95 |
|
|
Sapling |
Unidentified species 1 local (Jampu-jampu) |
59.22 |
|
Shorea koordesii |
37.43 |
|
|
Knema tomentella |
10.02 |
|
|
Dillenia serrata |
7.44 |
|
|
Unidentified species 1 local; (Awo-awo) |
7.13 |
|
|
Diospyros celebica |
6.80 |
|
|
Seedling |
Endiandra rubescens |
38.90 |
|
Amorphophallus konjac |
24.69 |
|
|
Intsia bijuga |
19.73 |
|
|
Calophyllum spectabile |
19.27 |
|
|
Vitex cofassus |
15.53 |
|
|
Shorea koordesii |
13.42 |
Table 5. Dominant species based on the important value index (IVI) in the natural D. celebica community at Maros
|
Growth Stratum |
Species |
IVI |
|
Tree |
Diospyros celebica |
119.07 |
|
Arenga pinnata |
66.73 |
|
|
Mangifera quadrifida |
24.74 |
|
|
Aleurites moluccana |
20.36 |
|
|
Myristica impressa |
8.85 |
|
|
Palaquium obovatum |
7.02 |
|
|
Pole |
Diospyros celebica |
129.80 |
|
Areca catechu |
60.54 |
|
|
Garcinia bancana |
13.69 |
|
|
Cinnamomum iners |
13.59 |
|
|
Mangifera quadrifida |
11.31 |
|
|
Sapling |
Diospyros celebica |
70.84 |
|
Cinnamomum iners |
46.67 |
|
|
Areca catechu |
37.51 |
|
|
Dracontomelon dao |
18.55 |
|
|
Leea indica |
12.93 |
|
|
Garcinia bancana |
12.79 |
|
|
Seedling |
Diospyros celebica |
31.29 |
|
Palaquium obovatum |
16.63 |
|
|
Areca catechu |
14.53 |
|
|
Cinnamomum iners |
12.00 |
|
|
Arenga pinnata |
11.69 |
|
|
Psychotria sp. |
10.26 |
3.5 Species associations in ebony communities
Significant species associations involving D. celebica varied among sites and growth strata (Table 6). In Maros, ebony showed a positive association with Cinnamomum iners and Palaquium obovatum in the sapling and seedling layer, as indicated by a moderate Ochiai index, suggesting that both species tend to share similar microhabitats and can coexist in the subcanopy. By contrast, ebony exhibited negative associations with Arenga pinnata in the tree and seedling layers, which may reflect competition for light and space, particularly during early regeneration.
Table 6. Significant associations between D. celebica and other species in the natural ebony communities at Maros, Sidrap, and Barru
|
Site |
Stratum |
Associated Species |
Association Type (χ²) |
Ochiai Index (IO) |
Strength Category |
|
Maros |
Tree |
Arenga pinnata |
Negative |
0.25 |
Low |
|
Sapling |
Cinnamomum iners |
Positive |
0.42 |
Medium |
|
|
Seedling |
Palaquium obovatum |
Positive |
0.35 |
Medium |
|
|
Barru |
Pole |
Knema pubiflora |
Positive |
0.55 |
High |
|
Sapling |
Lasianthus firmus |
Positive |
0.40 |
Medium |
|
|
Seedling |
Arenga pinnata |
Negative |
0.30 |
Low |
|
|
Sidrap |
Tree |
Payena acuminata |
Positive |
0.38 |
Medium |
|
Sapling |
Shorea koordesii |
Positive |
0.33 |
Medium |
In Barru, a strong positive association was detected between D. celebica and Knema pubiflora in the pole layer, indicating that these species frequently occur together in relatively protected microsites that favour intermediate growth stages. Ebony also showed positive associations with some sapling species, such as Lasianthus firmus, reinforcing the idea of species groupings that characterize the mid-storey of the stand. Conversely, negative associations with dominant seedling-layer species like Arenga pinnata are consistent with the low IVI of ebony in this stratum and point to intense competition at the seedling stage.
In Sidrap, associations between ebony and other species were generally weaker, with a few positive relationships detected in the tree and sapling layers, for example, with Payena acuminata and Shorea koordesii. These patterns suggest that, although D. celebica still structures the current canopy, its regeneration is less tightly linked to specific companion species and more exposed to competition from a broader set of taxa. Taken together, the association patterns across sites highlight that the persistence of ebony depends not only on its own demographic structure but also on its interactions with co-occurring species, especially in the regeneration strata.
3.6 Ecology indices
Table 7 shows that vegetation community structure exhibited strong differentiation across sites and growth stages, reflecting variations in site conditions and successional dynamics. Sidrap exhibited the highest community complexity in the mature strata, with high Shannon–Wiener diversity values at the tree (H′ = 3.35) and pole (H′ = 3.38) levels and very high evenness (J′ = 0.85–0.87), indicating a relatively balanced distribution of individuals among species and a stable stand structure (Table 7). In contrast, Maros and Barru had lower diversity and evenness in the tree and pole strata (H′ = 2.14–2.63; J′ = 0.53–0.70), indicating the dominance of certain species and a simplification of the community structure in the mature stage. In regeneration strata, Barru was characterized by the highest diversity and evenness at the sapling stage (H′ = 3.42; J′ = 0.86), while Maros showed the highest seedling diversity (H′ = 3.00; J′ = 0.79), indicating relatively diverse initial recruitment processes but with different regeneration trajectories between sites. Species richness patterns reinforced these findings, with Barru having the highest tree richness (DMg = 8.90) while Sidrap maintained relatively high and consistent richness in both tree and pole strata (DMg = 6.17–6.65). Analytically, this combination of diversity, evenness, and richness values suggests that community structure at each site is shaped by interactions between edaphic conditions, disturbance levels, and successional processes, resulting in high community stability in Sidrap, structural dominance in Maros, and dynamic but inhibited initial regeneration towards maturity strata in Barru.
Table 7. Ecology index in the natural D. celebica communities at Maros, Sidrap, and Barru
|
Index |
Growth Stage |
Sidrap |
Maros |
Barru |
|
Shannon–Wiener diversity (H′) |
Tree |
3.35 |
2.15 |
2.14 |
|
Pole |
3.38 |
1.81 |
2.63 |
|
|
Sapling |
2.98 |
2.62 |
3.42 |
|
|
Seedling |
2.27 |
3.00 |
2.91 |
|
|
Margalef richness (DMg) |
Tree |
6.17 |
4.82 |
8.90 |
|
Pole |
6.65 |
3.58 |
5.70 |
|
|
Sapling |
4.14 |
4.09 |
4.96 |
|
|
Seedling |
2.91 |
4.24 |
3.10 |
|
|
Pielou’s evenness (J′) |
Tree |
0.85 |
0.61 |
0.53 |
|
Pole |
0.87 |
0.54 |
0.70 |
|
|
Sapling |
0.83 |
0.72 |
0.86 |
|
|
Seedling |
0.80 |
0.79 |
0.81 |
3.7 Principal component analysis (PCA)
3.7.1 Population of D. celebica
PCA analysis showed that the population structure of Diospyros celebica was closely related to the soil property gradient shown in Figure 5. Seedling, sapling, and tree densities were positively correlated with the first Principal Component (PC1), which was characterized by high organic C, CEC, porosity, pH, and phosphorus availability, indicating that ebony regeneration and adult presence were better in soils with relatively fertile chemical and physical conditions. In contrast, fine-textured fractions (clay and silt) and total N were negatively associated with PC1, indicating that the dominance of these fractions did not always support ebony regeneration. Pole-level density mainly contributed to the second Principal Component (PC2), indicating a population response at the intermediate growth stage that is more sensitive to variations in site conditions.
3.7.2 Population of the D. celebica ecosystem
PCA indicated that overall variation in vegetation community structure was primarily driven by edaphic gradients shown in Figure 6. PC1 reflected gradients in soil fertility and physical quality, with strong positive loadings on organic carbon, CEC, porosity, pH, available phosphorus, and sand fractions, which correlated with increasing individual densities across all growth strata. Conversely, fine texture (clay and silt) and total nitrogen fractions had negative loadings on PC1, indicating limited community development in denser, less aerated soils. PC2 represented stand structural variation, primarily influenced by pole- and tree-stage densities, reflecting differences in successional dynamics and community responses to heterogeneity in site conditions.

Figure 5. PCA of the relationship between the population structure of Diospyros celebica and soil properties

Figure 6. PCA of the relationship between the population structure of the Diospyros celebica community and soil properties
The contrasting population structures and community patterns of Diospyros celebica observed among the three sites indicate that natural ebony ecosystems in South Sulawesi are at different ecological stages and face distinct regeneration challenges. In Maros, the high density of individuals across all growth stages and the strong dominance of ebony in the tree and pole layers reflect a population structure approaching a reverse J-shaped distribution, which is widely recognized as an indicator of healthy and continuous regeneration in tropical forests. Similar inverted J-shaped or exponentially declining size-class curves have been interpreted as signs of good regeneration and stable population dynamics in other tropical tree species and forest types [20, 21]. By contrast, the patterns in Barru and especially Sidrap deviate from this ideal, suggesting that regeneration of ebony is either constrained at specific life stages or increasingly outcompeted by other species [22].
The Barru site illustrates a typical “bottlenecked” regeneration pathway: Ebony shows very high importance in the pole layer and remains ecologically relevant in the sapling layer, yet its importance in the seedling layer is extremely low compared to understory palms and herbs [23, 24]. This pattern suggests that, although many individuals survive to intermediate stages, the earliest regeneration phase is strongly suppressed, probably due to edaphic limitations [25] combined with intense competition for light and space in the understorey [26]. Soil data show that Barru has the lowest pH values among the three sites, implying more acidic conditions that can reduce nutrient availability and constrain early seedling growth [27, 28]. Studies on ebony growing media and soil amendments suggest that sub-optimal chemical properties can significantly affect juvenile performance and thus long-term regeneration potential [29]. These findings are consistent with research on other tropical forests where poor regeneration status is often associated with interrupted or distorted reverse J-shaped curves, reflecting high mortality or recruitment failure in specific size or age classes [26, 30].
In Sidrap, ebony still dominates the tree layer but exhibits low importance in the pole, sapling, and seedling layers, where other species such as Payena acuminata, Shorea koordesii, and several understorey taxa become more prominent [31]. This pattern suggests that the current dominance of ebony in the canopy is a legacy of past conditions, while the regeneration layer is already shifting towards a different species composition. Similar situations, where high-value tree species maintain adult dominance but show poor recruitment, have been documented in other tropical regions and interpreted as symptoms of “demographic erosion” that may lead to future loss of these species from the canopy if no management intervention occurs [32, 33]. For ebony, such a trend is particularly concerning given its high economic value, slow growth, and Endangered conservation status, which together imply that recovery from recruitment failure will be slow and uncertain [13].
The strong dominance of D. celebica in Maros contrasts sharply with earlier reports from Central Sulawesi [13], where regeneration density of ebony seedlings and saplings was relatively low and often insufficient to compensate for ongoing disturbances. In the Maleali seed stand, for example, ebony regeneration was described as limited compared with other sites, despite the area being designated for seed production [13]. Other studies on ebony in Sulawesi also highlight that juvenile stages under the canopy of mother trees are often sparse and vulnerable to disturbance. In this context, the Maros population, with its high importance values across multiple strata, appears to represent a comparatively robust natural stronghold for ebony in South Sulawesi. However, the presence of substantial competition from co-dominant species in the sapling and seedling layers indicates that this stability is not guaranteed and may deteriorate if disturbance intensity increases [17, 34].
The species association patterns further illuminate the ecological interactions shaping ebony communities at each site [33]. Positive associations between D. celebica and species such as Cinnamomum iners, Knema pubiflora, or Payena acuminata suggest that these taxa share similar microhabitats and may form relatively stable species assemblages in the subcanopy and mid-storey. Such co-occurring species groups have been reported in other tropical forests and are often linked to specific combinations of soil conditions, canopy structure, and disturbance regimes. In contrast, negative associations between ebony and understorey dominants such as Arenga pinnata at the seedling layer indicate strong competitive interactions that can inhibit early regeneration of ebony. Studies on sugar palm and similar Arecaceae-dominated systems have shown that dense palm understorey can substantially modify light conditions and resource availability [35], thereby altering tree regeneration patterns and overall community composition [6]. The association results from Barru and Sidrap are consistent with this, supporting the interpretation that understorey competition is a key factor limiting ebony recruitment at some sites.
From a conservation and management perspective, these findings underscore the need for site-specific strategies to sustain ebony populations in South Sulawesi. In Maros, management should prioritize the strict protection of the existing mature stands and maintenance of habitat quality to preserve the currently favourable regeneration dynamics. In Barru, interventions should focus on enhancing early regeneration, for instance by locally reducing competitive understorey cover and improving microsite conditions for seedling establishment. In Sidrap, where regeneration is already dominated by non-ebony species, more intensive measures such as enrichment planting, assisted natural regeneration, or targeted control of aggressive competitors may be required to prevent further decline of ebony in the canopy. Comparable recommendations have been proposed for other tropical tree species showing poor regeneration, where assisted natural regeneration and focused management of critical life stages were found to improve population viability [22, 36, 37].
Finally, when viewed alongside previous work on the autecology, genetic mating system, and reproductive biology of ebony, the present study fills an important gap by providing a detailed picture of population structure and community context across multiple natural habitats [8, 38, 39]. High outcrossing rates and effective pollen dispersal have been reported for D. celebica, indicating that, from a reproductive biology standpoint, the species has the potential to maintain genetic connectivity across stands [18]. Thus, the main constraints to its persistence at the landscape scale are likely to be habitat degradation, overexploitation of large trees, and failures in early regeneration driven by competition and suboptimal edaphic conditions rather than intrinsic reproductive limitations. This highlights the urgency of integrating stand-level vegetation management with broader landscape-scale conservation planning to secure the long-term future of ebony in Sulawesi’s tropical forests.
Natural ebony ecosystems in South Sulawesi represent distinct demographic states along ecological gradients, with stable, self-replacing populations in Maros, a bottlenecked stand in Barru, and an aging, demographically eroding stand in Sidrap. These differences appear closely linked to site-specific edaphic conditions and to species interactions within the understorey. More acidic soils and the strong dominance of shade-tolerant palms and herbs in Barru appear to limit the establishment and survival of ebony seedlings, while in Sidrap, the regeneration layer is increasingly occupied by co-occurring tree species that compete effectively with ebony for light and space. Such patterns indicate that, beyond harvesting pressure, local habitat quality and biotic competition are key determinants of ebony regeneration success and long-term persistence in these natural forests.
Practically, this implies that the findings highlight the need for differentiated, site-based conservation strategies. In Maros, priority should be given to the strict protection of mature stands and the maintenance of favourable habitat conditions to sustain the currently robust regeneration dynamics. In Barru, management efforts should focus on improving the performance of early life stages, for example, by reducing competitive understorey cover and enhancing microsite conditions for seedling establishment. In Sidrap, where regeneration is already strongly dominated by non-ebony species, more intensive measures such as enrichment planting and assisted natural regeneration will be required to secure the future role of ebony in the canopy.
Overall, this study underscores the value of integrating population structure, important value indices, soil properties, and species association analyses to diagnose the ecological status of endangered tree species and their habitats. Such an integrated approach provides a robust scientific basis for designing targeted, evidence-based interventions to conserve Diospyros celebica across its remaining natural range and contributes to broader efforts to maintain the integrity and resilience of tropical forest ecosystems in Sulawesi.
This research was conducted based on the Decree of the National Research and Innovation Agency (BRIN), Number 37/II.7/HK/2023, concerning Recipients of the Research and Innovation Program for Advanced Indonesia Batch-4. Furthermore, a Cooperation Agreement was signed between BRIN and the Institute for Research and Community Service (LPPM) of Hasanuddin University, Number 184/IV/KS/11/2023. We, the authors, would like to thank BRIN and LPDP RI for funding this research, which made it possible to conduct the research successfully. We also thank LPPM-Hasanuddin University for their cooperation in facilitating the administrative process.
[1] Artaxo, P., Hansson, H.C., Machado, L.A.T., Rizzo, L.V. (2022). Tropical forests are crucial in regulating the climate on Earth. PLOS Climate, 1(8): e0000054. https://doi.org/10.1371/journal.pclm.0000054
[2] Dicky, D., Paembonan, S.A., Mukrimin, M., Millang, S., Witno, W., Makkasau, A.R., Arty, B., Ramadhan, W., Rasyid, A. (2025). Comparison of carbon storage and oxygen release in various land uses in North Luwu District, South Sulawesi Province, Indonesia. Biodiversitas, 26(6): 2754-2761. https://doi.org/10.13057/biodiv/d260620
[3] Zheng, B.F., Wang, D., Chen, Y.X., Jiang, Y.H., Hu, F.Q., Xu, L.L., Zhang, J.H., Zhu, J.Q. (2024). The impact of vegetation types on soil hydrological and mechanical properties in the hilly regions of Southern China: A comparative analysis. Water, 16(2): 350. https://doi.org/10.3390/w16020350
[4] Aldiansyah, S., Risna, R. (2023). Assessing potential habitat suitability of vulnerable endemic species: A case study of Diospyros celebica Bakh and Rhyticeros cassidix. Forum Geografic, XXII(2): 159-169. https://doi.org/10.5775/fg.2023.2.3596
[5] Kuswandi, R., Murdjoko, A. (2015). Population structures of four tree species in logged-over tropical forest in South Papua, Indonesia: An integral projection model approach. Indonesian Journal of Forestry Research, 2(2): 93-101. https://doi.org/10.59465/ijfr.2015.2.2.93-101
[6] Germain, S.J., Hansen, W.D., Canham, C.D. (2025). Reassembly of disturbed forests portends climate resilience but diversity loss. Ecosystems, 28: 9. https://doi.org/10.1007/s10021-024-00959-0
[7] Fatlan, K.F., Pamoengkas, P., Majiidu, M., Siregar, I.Z. (2021). Tree species diversity on ebony habitat with different degradation levels in Sulawesi. IOP Conference Series: Earth and Environmental Science, 918: 012052. https://doi.org/10.1088/1755-1315/918/1/012052
[8] Jakovac, C.C., Junqueira, A.B., Crouzeilles, R., Peña-claros, M., Mesquita, R.C.G., Bongers, F. (2021). The role of land-use history in driving successional pathways and its implications for the restoration of tropical forests. Biological Reviews, 96(4): 1114-1134. https://doi.org/10.1111/brv.12694
[9] Nurkin, B., Allo, M.K, Gintings, A.N. (2018). Eboni Sulawesi (Diospyros celebica Bakh.). UPT Unhas Press.
[10] Asdar, M., Prayitno, T.A., Lukmandaru, G., Faridah, E. (2015). Distribution, potency and quality ebony (Diospyros celebica Bakh.) in Sulawesi. Jurnal Ilmu-ilmu Pertanian, 22(2): 94-105.
[11] Martha, R., Mubarok, M., Darmawan, W., Syafii, W., Dumarcay, S., Charbonnier, C.G., Gérardin, P. (2021). Biomolecules of interest present in the main industrial wood species used in Indonesia-A review. Journal of Renewable Materials, 9(3): 399-449. https://doi.org/10.32604/jrm.2021.014286
[12] Kinho, J. (2013). Mengembalikan kejayaan eboni makasar di Sulawesi Utara [Restoring the glory of makasar ebony in North Sulawesi]. Forestry Research and Development Agency. Forestry Research Center Manado.
[13] Rukmi, R., Labiro, E., Rosyid, A., Yusran, Y., Mowidu, I. (2023). Potential, distribution pattern and autecology of Diospyros celebica Bakh. in the maleali seed stand area, Central Sulawesi, Indonesia. International Journal of Design & Nature and Ecodynamics, 18(2): 421-428. https://doi.org/10.18280/ijdne.180220
[14] Kusumadewi, Y., Wardani, W., Sudarmonowati, E., Partomihardjo, T., et al. (2020). Indonesian red list of plants 1: 50 species of commercial timber trees. https://doi.org/10.14203/press.310
[15] Suryawan, A., Kinho, J., Mayasari, A. (2011). Potensi permudaan alami jenis-jenis eboni (Diospyros spp.) di Cagar Alam Tangkoko, Bitung Sulawesi Utara. Info BPK Manado, 1(1): 21-34.
[16] Nurkin, B., Achmad, A., Oka, N.P., Rachman, W., Paembonan, S.A. (2002). Karakteristik ekologi dan aspek silvikultur Eboni (Diospyros celebica Bakh) Sulawesi Selatan [Ecological characteristics and silvicultural aspects of Ebony (Diospyros celebica Bakh) South Sulawesi]. Berila Biologi, 6(2): 267-275. https://media.neliti.com/media/publications/63219-ID-none.pdf.
[17] Saleha, S., Ngakan, P.O. (2016). Distribution and population structure of the juvenile of Diospyros celebica Bakh. under the canopy of their mother-tree. Jurnal Penelitian Kehutanan Wallacea, 5(2): 103-111. http://doi.org/10.18330/jwallacea.2016.vol5iss2pp103-111
[18] Restu, M., G., G., Larekeng, S.H. (2017). High outcrossing rate and pollen dispersal distance of Diospyros celebica Bakh. (Ebenaceae), an endemic tree species in Sulawesi Island, Indonesia. Biotropia, 24(3): 173-181. https://doi.org/10.11598/btb.2017.24.3.562
[19] Nuryanti, D.M., Liana, Witno, Soma, A.S., Ayuwidia, S.M., Awalia, I.M. (2023). Composition, diversity and association of pioneer plants on post- landslide areas in Palopo, South Sulawesi, Indonesia. Biodiversitas, 24(3): 1862-1870. https://doi.org/10.13057/biodiv/d240362
[20] Jubair, A.N.M., Rahman, M.S., Sarmin, I.J., Raihan, A. (2023). Tree diversity and regeneration dynamics toward forest conservation and environmental sustainability: A case study from Nawabganj Sal Forest, Bangladesh. Journal of Agriculture Sustainability and Environment, 2(2): 1-22. https://doi.org/10.56556/jase.v2i2.780
[21] Sarkar, M., Devi, A. (2014). Assessment of diversity, population structure and regeneration status of tree species in Hollongapar Gibbon Wildlife Sanctuary, Assam, Northeast India. Tropical Plant Research, 1(2): 26-36. https://www.tropicalplantresearch.com/archives/2014/vol1issue2/5.pdf.
[22] Marwa, J., Ungirwalu, A., Imburi, C.S., Djitmau, D.A., Murdjoko, A., Benu, N.M.H. (2024). Ecological perspective to sustainably manage the secondary forest in the lowland of Doberai Peninsula, Indonesia. Biodiversitas, 25(8): 2720-2732. https://doi.org/10.13057/biodiv/d250845
[23] Trauernicht, C., Ticktin, T. (2005). The effects of non-timber forest product cultivation on the plant community structure and composition of a humid tropical forest in southern Mexico. Forest Ecology and Management, 219(2-3): 269-278. https://doi.org/10.1016/j.foreco.2005.09.005
[24] Hernández-Salmerón, I.R., Delconte, V., van Hoeflaken, J., Solorio-Sánchez, F.J., Holmgren, M. (2023). Native palms and trees mediate drought impacts on dry neotropical pastures. Journal of Applied Ecology, 60(7): 1376-1387. https://doi.org/10.1111/1365-2664.14425
[25] Rahman, A., Monde, A., Widjajanto, D., Khaliq, M.A., Adrianton, Oktavina, T. (2024). Swallow droppings for the chemical properties of alluvial soil as growing medium for ebony (Diospyros celebica). Jurnal Penelitian Pendidikan IPA, 10(8): 5935-5942. https://doi.org/10.29303/jppipa.v10i8.7856
[26] Kueffer, C., Schumacher, E., Fleischmann, K., Edwards, P.J., Dietz, H. (2007). Strong below-ground competition shapes tree regeneration in invasive Cinnamomum verum forests. Journal of Ecology, 95(2): 273-282. https://doi.org/10.1111/j.1365-2745.2007.01213.x
[27] Kuśmierz, S., Skowrońska, M., Tkaczyk, P., Lipiński, W., Mielniczuk, J. (2023). Soil organic carbon and mineral nitrogen contents in soils as affected by their pH, texture and fertilization. Agronomy, 13(1): 267. https://doi.org/10.3390/agronomy13010267
[28] Khairunnisa, P., Bakri, B. (2025). Some chemical properties of mineral soil under peat layer at Sriwijaya Botanical Garden. Journal of Suboptimal Lands, 14(2): 154-160. https://doi.org/10.36706/JLSO.14.2.2025.690
[29] Mukrimin, M. (2023). Study of soil physical and chemical properties under ebony (Diospyros celebica Bakh.) stands in provenance of the Amaro protected forest, South Sulawesi. AIP Conference Proceedings, 2596: 060011. https://doi.org/10.1063/5.0118848
[30] Singh, S., Dixit, B., Prajapati, L., Chandrakar, S., Tamrakar, A. (2024). Characterization of species structure and regeneration patterns under different density gradients in a tropical Sal forest of AchanakmarAmarkantak biosphere reserve in Central India. Environmental Conservation Journal, 25(3): 824-835. https://doi.org/10.36953/ECJ.27542830
[31] Baig, M.A., Zehra, N., Khan, J.A. (2025). Population structure and regeneration status of tree species: An assessment in the ranikhet forest of Almora District, Kumaon Himalaya, Uttarakhand, India. International Journal of Ecology and Environmental Sciences, 51(3): 311-320. https://doi.org/10.55863/ijees.2025.0636
[32] Ao, A., Changkija, S., Tripathi, S.K. (2024). Patterns of forest community diversity, regeneration potential and carbon storages along an altitudinal gradient in Eastern Himalaya, India. Environmental and Sustainability Indicators, 22: 100399. https://doi.org/10.1016/j.indic.2024.100399
[33] Martini, F., Buechling, A., Bače, R., Hofmeister, J., Janda, P., Matula, R., Svoboda, M. (2025). Biotic and abiotic effects on tree regeneration vary by life stage in European primary forests. Oikos, 2025(2): e10755. https://doi.org/10.1111/oik.10755
[34] Lahiri, S., Dash, S.S. (2021). Community structure and regeneration status of tree species in kyongnosla alpine sanctuary, eastern Himalaya, India. Indonesian Journal of Forestry Research, 8(2): 241-257. https://doi.org/10.20886/ijfr.2021.8.2.241-257
[35] Kadir, Z.S., Baderan, D.W.K., Hamidun, M.S. (2025). Composition of species, utilization, and conservation status of plant species in the sugar palm (Arenga pinnata) agroforestry system. Social, Agricultural, Food System and Environmental Sustainability, 2(1): 1-16. https://doi.org/10.61511/safses.v2i1.2025.1810
[36] Gonçalves, A.C., Fonseca, T.F. (2023). Influence management and disturbances on the regeneration of forest stands. Frontiers in Forests and Global Change, 6: 1123215. https://doi.org/10.3389/ffgc.2023.1123215
[37] Joshi, N., Joshi, N. (2025). Comparative study of Sal (Shorea robusta Gaertn. f.) regeneration in assisted and non- assisted natural regeneration forests of Kumaun Himalaya, India. Current World Environment, 20(2): 698-704. https://doi.org/10.12944/CWE.20.2.13
[38] Damaševičius, R., Maskeliūnas, R. (2025). Modeling forest regeneration dynamics: Estimating regeneration, growth, and mortality rates in lithuanian forests. Forests, 16(2): 192. https://doi.org/10.3390/f16020192
[39] Henry, C.R., Walters, M.B. (2023). Tree species size class patterns portend compositional shifts and low resilience in managed northern hardwood forests. Ecosphere, 14(7): e4621. https://doi.org/10.1002/ecs2.4621