Effects of Potassium and Arbuscular Mycorrhiza on Yield, Starch, and Beta-Carotene of Local Sweet Potato (Ipomoea batatas L.)

Mardhiah Hayati*![]() | Hifnalisa Hifnalisa
| Hifnalisa Hifnalisa![]() | Murna Muzaifa
| Murna Muzaifa![]() | Cut Nur Ichsan
| Cut Nur Ichsan![]() | Gina Erida
| Gina Erida![]() | Nura Nura
| Nura Nura![]()
© 2025 The authors. This article is published by IIETA and is licensed under the CC BY 4.0 license (http://creativecommons.org/licenses/by/4.0/).
OPEN ACCESS
Improving sweet potato productivity, starch deposition, and β-carotene levels in nutrient-poor soils requires the synergistic use of potassium fertilization and Arbuscular Mycorrhizal Fungi (AMF) inoculation. This study investigated the optimal application rates of potassium and AMF and their interactive effects on growth performance, yield, starch accumulation, and β-carotene concentration in local sweet potato (Ipomoea batatas L.) clones from Saree, Aceh Province, Indonesia. A field experiment was conducted from April to September 2023 using a Randomized Block Design in a 4 × 3 factorial arrangement, comprising four potassium fertilizer (K₂O) rates (0, 60, 120, and 180 kg ha⁻¹) and three AMF application rates (0, 10, and 20 g plant⁻¹), with three replications. The findings of this study revealed that the highest yield, starch, and β-carotene content were achieved with a K2O dose of 120 kg ha-¹ (8.39 tons ha-1, 66.42%, 3.32%) and an AMF dose of 10 g plant-¹ (8.91 tons ha-1, 66.70%, 2.60%). Potassium fertilization enhanced sweet potato quality, with peak starch content (69.36%) at 60 kg ha⁻¹ K₂O and highest harvest index (0.77) and β-carotene (3.37%) at 120 kg ha⁻¹ K₂O, both without AMF. While potassium markedly improved yield and nutritional traits, AMF showed no consistent quality benefits or synergy with potassium under these conditions.
arbuscular mycorrhiza, β-carotene, chlorophyll, harvest index, local clones, starch
Sweet potato (Ipomoea batatas L.) is a key food crop in tropical and subtropical regions, providing an important source of human nutrition, animal feed, functional food ingredients, and raw materials for the food industry [1]. This crop is known for its high adaptability to suboptimal environments, tolerance to drought, and its nutritional content, which includes carbohydrates (20%), fiber, vitamins A and C, anthocyanins, starch, and β-carotene, which are very important in efforts to overcome vitamin A deficiencies such as night blindness and xerophthalmia [2]. With a starch content of 33-64% dry weight, sweet potato serves as a versatile ingredient for food diversification, wheat flour replacement, and climate-adaptive formulations such as emergency biscuits.
Enhancing sweet potato production remains a critical research priority, particularly in developing countries where the crop constitutes a major component of food security. Despite its global significance, Indonesia experienced a 5.34% decline in national sweet potato output in 2023, decreasing from 1.51 million tons to 1.43 million tons [3]. Sweet potato cultivation in Indonesia is limited by poor soil fertility, low-input, local varieties, and minimal use of modern agronomic practices. Saree clones grown on unfertilized marginal soils showed low carbohydrate content (12.89%), small tuber size, and modest yields (10.83 tons ha-1) [4, 5]. These findings support a strategy to improve yield and quality in low-input systems.
Potassium (K) is a vital macronutrient that supports key physiological processes, including photosynthesis, assimilate transport, root elongation, stomatal regulation, stress tolerance, protein synthesis, and enzyme activation. In addition, K plays a particularly important role in tuber development and sugar accumulation [6]. It has shown significant effects on sweet potato growth and yield across varieties. The Sari variety achieved optimal performance with 160 kg ha⁻¹ KCl, yielding 35.59 tons ha-1 [7], while Narutokintoki responded to 120-125.5 kg ha-1 K₂O with yield increases up to 21 tons ha-1 [8]. Potassium also enhances starch biosynthesis and tuber formation by regulating key enzymes (SuSy, AGPase, and SSS) involved in sucrose conversion [9]. Additionally, co-application of 150 kg ha-1 K₂O with 30 ppm Zn significantly improved tuber carotene content [10].
Biofertilizers such as Arbuscular Mycorrhizal Fungi (AMF), particularly Glomeromycota, enhance nutrient uptake, root architecture, and yield under low-fertility conditions [11, 12]. Through mutualistic symbiosis, AMF form hyphal networks that mobilize subsoil nutrients, especially phosphate, making them more accessible to plant roots [11]. AMF also contribute to increased β-carotene content, particularly under low-phosphorus conditions, indicating their role in stimulating secondary metabolism [13]. Sweet potato is typically harvested at five months, though maturity varies by environment and cultivar [14]. β-carotene levels are influenced by agro-ecological factors, temperature, tuber maturity, harvest timing, and postharvest storage duration [15, 16].
AMF act as biostimulants by enhancing organic matter decomposition and nutrient mineralization, thereby improving plant growth in low-fertility soils [12]. Their mutualistic association with roots facilitates phosphorus uptake via subsoil hyphal networks, contributing to sustainable agriculture by reducing dependence on synthetic inputs [17]. In sweet potato, application of 25 g mixed AMF increased branching, tuber biomass, and overall yield by week 20, with further gains observed when combined with N and K fertilizers, highlighting the importance of early nutrient availability for effective root colonization and hyphal development [18].
Despite growing evidence of the benefits of AMF in enhancing sweet potato growth and quality, most studies have focused on common varieties and single-factor applications. Potassium, a key nutrient in photosynthate transport, starch biosynthesis, and β-carotene accumulation, remains underexplored in combination with AMF, particularly in local clones under specific agroecological conditions. This study investigates the interactive effects of potassium dosage and AMF application on the growth, yield, starch content, and β-carotene concentration of Saree local sweet potato clones, aiming to identify optimal nutrient combinations for improved agronomic and nutritional performance.
2.1 Place and time of research
The study was conducted from April to September 2023 in Saree, Aceh Province, Indonesia, which is located at a moderate elevation of 619 m above sea level (at coordinates 95°42'17 E and 5°26'5 N). The area is classified as climate type C (moderately wet) based on the Schmidt and Ferguson classification. The analysis of AMF colonization was carried out at the Plant Physiology Laboratory, Faculty of Agriculture, Universitas Syiah Kuala. Soil analysis was conducted at the Soil, Plant, Fertilizer, Water Laboratory of the Spice and Medicinal Plants Research Institute, Cimanggu, Bogor. The starch content was measured at the Laboratory of the Agricultural Equipment Standardization Center, Ministry of Agriculture. Climatic data during the research period are presented in Table 1. Table 1 presents the average rainfall and temperature during the study period. Vegetative and generative period observations were conducted separately, with data collected in April–May and June–September, respectively.
Table 1. Climatic data (rainfall, average air temperature, daily maximum and minimum temperature, and air humidity) from April to September 2023
|
Period |
Month |
Rainfall (mm) |
Average Air Temperature (℃) |
T max (℃) |
T min (℃) |
RH (%) |
|
Vegetative |
April |
36.50 |
27.20 |
34.57 |
22.10 |
81.67 |
|
May |
45.50 |
27.77 |
35.17 |
23.07 |
78.67 |
|
|
Average |
41.00 |
27.48 |
34.87 |
22.58 |
80.17 |
|
|
Generative |
June |
7.03 |
28.10 |
36.03 |
22.20 |
73.67 |
|
July |
15.80 |
28.17 |
35.10 |
22.70 |
72.67 |
|
|
August |
28.23 |
27.43 |
34.93 |
22.83 |
77.00 |
|
|
September |
64.95 |
27.20 |
35.20 |
22.50 |
80.00 |
|
|
Average |
29.00 |
27.73 |
35.32 |
22.56 |
75.83 |
Source: Aceh Climatology Station, Meteorology, Climatology and Geophysics Agency, Indrapuri Meteorological Station, Aceh Besar, Indonesia.
2.2 Research materials and tools
The plant material used in this study consisted of shoot cuttings from local sweet potato clones of Saree, each measuring 30 cm in length. Additional materials included plastic bags, decomposed cow manure applied at a rate of 10 tons ha⁻¹, potassium fertilizer (according to treatment), urea (90 kg N ha⁻¹), SP-36 (75 kg P₂O₅ ha⁻¹), and AMF biofertilizer (a mixture of Glomus sp. and Gigaspora sp. in a 1:1 ratio with added zeolite), applied according to the treatment. The equipment used comprised an oven, analytical balance, mortar and pestle, 80-mesh sieve, SPAD chlorophyll meter, and spectrophotometer.
2.3 Research design and data analysis
This study used a Randomized Block Design (RBD) with a 4 × 3 factorial arrangement. The first factor was the potassium fertilizer (K₂O) dose at four levels (0, 60, 120, and 180 kg ha⁻¹), and the second factor was the AMF dose at three levels (0, 10, and 20 g plant⁻¹). Each treatment combination was replicated three times. AMF colonization was evaluated six weeks after planting (WAP), while chemical analyses of starch and β-carotene content, along with tuber yield assessment, were conducted at 17 WAP. Data were analyzed using an F-test, followed by Tukey’s Honestly Significant Difference (HSD) test at the 5% significance level. All statistical analyses were carried out using SPSS version 22.
2.4 Soil analysis
Prior to the experiment, soil samples were randomly collected from several locations within the study area at a depth of 0-20 cm. The samples were analyzed to assess soil nutrient status and fertility. The results showed a soil pH of 4.80, C-organic content of 3.36%, total N of 0.52%, and a C/N ratio of 6.43. Available P₂O₅ was 0.53 ppm, while exchangeable bases were Ca 4.06 cmol kg⁻¹, Mg 0.32 cmol kg⁻¹, K 1.27 cmol kg⁻¹, and Na 0.29 cmol kg⁻¹, with a total base content of 5.90 cmol kg⁻¹. Exchangeable Al was 0.34 cmol kg⁻¹, and the soil cation exchange capacity reached 37.05 cmol kg⁻¹ with a base saturation of 15.93%. The soil texture consisted of 68.51% sand, 18.15% silt, and 13.34% clay. According to soil fertility assessment criteria, the soil at the study site is classified as low in fertility.
2.5 Sweet potato planting, maintenance, and harvesting
Land preparation began with weed removal and soil loosening, followed by the construction of 36 ridges measuring 120 cm in width, 40 cm in height, and 60 cm in spacing between ridges. Each ridge was 500 cm long for each treatment unit. Sweet potato cuttings were planted at a spacing of 40 cm × 25 cm, with one cutting per hole. Organic fertilizer in the form of manure was applied at a rate of 10 tons ha⁻¹, one week prior to planting. Potassium and AMF fertilizers were applied at planting according to the assigned doses. AMF inoculum was placed in each planting hole before transplanting to ensure root contact, covered with soil to maintain spore viability, and watered, while potassium fertilizer was applied to the plant side. Nitrogen fertilizer (urea) was applied in three splits: one-third of the total dose at planting and the remaining two-thirds at two WAP. The full dose of phosphorus fertilizer (P₂O₅ from SP-36) was applied at planting. Irrigation was carried out as needed, while weed control was conducted every two weeks. Hilling and branch training were performed at 8, 12, and 15 WAP. Sweet potato tubers were harvested at 17 WAP for evaluation of yield and tuber quality.
2.6 Observed variables
The variables observed in the vegetative period were the number of branches and the length of branches plant-1 at the ages of 2, 4, and 6 WAP. The amount of chlorophyll, percentage of AMF colonization, weight of fresh biomass, root length, and root weight were observed at the age of 6 WAP. At the harvest age (17 WAP), which corresponds to the generative period of the plant, the following parameters were recorded: fresh and weight of dry biomass of the plant-1 (g) (obtained by oven-drying at 60℃ for 72 hours until a constant weight was achieved), the weight of tubers plant-1 (g); yield (ton ha-1), number of tubers-1 (tuber), tuber length (cm) and tuber diameter (mm), harvest index, starch (%) and β-carotene content (%) of sweet potato.
2.6.1 Chlorophyll content
Observation of chlorophyll content in leaves was carried out at the age of 6 WAP by using a chlorophyll meter (SPAD) with five samples of ridge plants-1. The sample taken was the 4th leaf from the top. The measured leaves were those that had opened completely. Observations were made by observing the amount of chlorophyll in three parts of the leaf (the base, middle, and tip) and then averaging.
2.6.2 Percentage of mycorrhizal colonization
After the plants were six weeks old, AMF-infected root samples were analyzed using a microscope with the following steps: roots were washed with distilled water until clean, then cut into 1 cm pieces and soaked in a 10% KOH solution for 24 hours to whiten the roots. Next, the roots were rinsed four times with water and then soaked in a 0.05% trypan blue solution and lactophenol for 24 hours. The roots were re-soaked using distilled water to clean them of the blue color. Finally, 10 roots were taken per plant, randomly placed on a glass preparation to be observed under a microscope (magnification 100-400 times), and later the percentage of infected/colonized roots was calculated using the following formula [19].
$\operatorname{AMF}$ colonization $(\%)=100 \times \frac{\text {number of infected roots}}{\text {number of roots observed}}$ (1)
2.6.3 Chemical analysis of starch content by the Luff-Schoorl method
Sweet potato tubers were thinly sliced, oven-dried at 105℃ for 24 hours, ground with a mortar, and sieved. A 5 g subsample was placed in a 500 mL Erlenmeyer flask and extracted with 200 mL of 3% HCl solution. The mixture was refluxed for three hours and, after cooling, neutralized with 30% NaOH, using litmus paper or phenolphthalein as an indicator. Subsequently, a small amount of 3% CH₃COOH was added to re-acidify the solution. The solution was transferred to a 500 mL volumetric flask. From the filtrate, 10 mL was pipetted into a 500 mL Erlenmeyer flask, to which 25 mL of Luff’s solution, a few boiling stones, and 15 mL of distilled water were added. The mixture was heated over a steady flame to reach a boil within three minutes and maintained at a boil for exactly 10 minutes, after which it was rapidly cooled in an ice bath. Following cooling, 15 mL of 20% KI solution and 25 mL of 25% H₂SO₄ were slowly added. Titration was then carried out immediately with 0.1 N Na₂S₂O₃ solution, with a blank sample prepared using the same procedure. The reducing sugar content was calculated using the formula: (Blank-Sample)×N×10, and the Luff-Schoorl table was used to determine the sugar concentration (mg) based on the sample volume (mL).
Glucose content $=\left(\frac{W \times d f}{W_1}\right) \times 100 \%$ (2)
where,
Starch content = 0.90×glucose content
W1 = Sample weight (mg)
W = Glucose contained for ml of sample used (mg)
df = Dilution factor
2.6.4 Chemical analysis of β-carotene by the thin-layer chromatography method
Approximately 0.25 g of the sweet potato sample was placed into a 25 mL volumetric flask, followed by adding chloroform to approximately one-quarter of the flask volume. The mixture was then shaken using a shaker for two hours. Afterward, the volume was adjusted to the mark and stored for 24 hours. The resulting concentrated extract was prepared at a specific concentration and analyzed using the Thin Layer Chromatography (TLC) method, with silica gel plates as the stationary phase. The extract was filtered, and 5 µL of the filtrate was applied to a silica gel 60 GF254 aluminum sheet plate. A β-carotene standard, dissolved in ethanol at a concentration of 400 ppm, was also applied at 5 µL. Elution was carried out using a chloroform, ethanol, and ethyl acetate solvent system until the solvent front reached approximately 15 cm. After elution, the TLC plate was dried and scanned using a Camag TLC Scanner 3 at a wavelength (λ) of 299 nm. The concentration was calculated using the following formula:
β-carotene content:
$\left(\frac{\left(\frac{\text {sample area}}{\text {standard area}}\right) \times(p p m \text { standard}) \times(\text {sample volume}) \times\left(\frac{d f(25000 \mu l)}{(5 \mu)}\right)}{m g \text { sample}}\right) \times 100 \%$ (3)
where, df = dilution factor.
2.6.5 Yield and harvest index
Yield (ton ha-1) was calculated based on the following formula:
Yield $=\left(\frac{\text {Land area}(\text {hectare})}{\text {Plant spacing}}\right) \times\left(\right.$ tuber weight plant $\left.^{-1}\right)$ (4)
Next, the harvest index (HI) is calculated using the following relationship.
Harvest Index $=\frac{\text {Total tuber yield}}{\text {Total Biomass Weight}+ \text {tubers}}$ (5)
3.1 Effect of potassium fertilizer dosage and mycorrhiza on sweet potato plant
3.1.1 Number of branches and branch length at 6 WAP
Sweet potato plants receiving 180 kg ha⁻¹ K₂O showed a non-significant trend toward higher branch numbers at 2-6 WAP, indicating potassium’s limited role in early branching compared with nitrogen. Potassium primarily supported photosynthetic activity and tuber development, with higher doses enhancing branch and leaf growth, chloroplast density, and biomass, consistent with previous reports [20, 21]. Increased leaf N content at 96 kg ha⁻¹ K₂O [22] suggests a possible K-N synergism in promoting vegetative and physiological performance. At 4 WAP, sweet potato plants receiving 60 kg ha⁻¹ K₂O exhibited greater branch length than the control, though differences from higher potassium treatments were not significant (Table 2). This rate was more effective than the control in promoting branch elongation but showed no clear advantage over higher doses. According to study [23], potassium enhances branch elongation, plant vigor, and leaf expansion during early growth, with growth parameters declining above 60 kg ha⁻¹, suggesting this may be an optimum level. In contrast, study [22] reported that 120 kg ha⁻¹ K₂O produced the tallest plants, significantly exceeding other treatments. The role of potassium in stimulating shoot growth is further supported by study [21], which observed significant increases in branch length, leaf number, and branch number per plant following potassium application. These effects are attributed to potassium’s role in enhancing photosynthesis, CO₂ assimilation, nitrogen metabolism, and translocation of photoassimilates [24]. Additionally, study [22] highlighted its role in stimulating enzyme activity, nutrient translocation, and protein biosynthesis, while study [21] found that plant height and biomass accumulation increased significantly with potassium rates from 40 to 120 kg ha⁻¹.
At 2 WAP, sweet potato plants treated with 20 g plant⁻¹ AMF showed a trend toward increased branching, though differences were not statistically significant (Table 2). This aligns with study [18], which reported no significant effect at early stages (4 WAP) but observed increased branching at 12 and 20 WAP with 25 g plant⁻¹ AMF. In the present study, branch numbers at 4 and 6 WAP remained higher under the 20 g plant⁻¹ AMF treatment, significantly exceeding the control but comparable to the 10 g dose. Similarly, study [25] found significant branching improvements with 10 g plant⁻¹ AMF. These effects likely stem from enhanced phosphate and nitrogen uptake facilitated by AMF hyphal networks, which promote nutrient mineralization and acquisition from organic nitrogen sources [26]. Table 2 shows that sweet potato plants treated with 20 g plant⁻¹ AMF had the longest branches at 2 and 6 WAP, although the difference from the 10 g plant⁻¹ treatment was not significant. Research [25] reported that branch length increased with AMF doses from 10 to 30 g plant⁻¹, likely due to the production of auxins and gibberellin-like substances by AMF [27]. Similarly, study [28] found that 20 g plant⁻¹ AMF enhanced branch length in sweet potato. These results suggest that AMF application improves nutrient availability. According to research [12], AMF also increases gibberellin levels, promoting stem elongation through enhanced cell division. Greater cell numbers accelerate stem growth, resulting in taller plants compared to uninoculated controls. Furthermore, AMF symbiosis improves water and phosphorus uptake, regulates plant hormones, and enhances tolerance to abiotic stress, collectively contributing to increased growth and productivity [27].
Table 2. Average number and length of sweet potato branches at 2, 4, and 6 WAP due to potassium fertilizer and AMF
|
K2O Dose (kg ha-1) |
Number of Branches (plant-1) |
Branch Length (cm) |
||||
|
2 WAP |
4 WAP |
6 WAP |
2 WAP |
4 WAP |
6 WAP |
|
|
0 |
1.47±0.43 a |
3.16±0.22 a |
3.26±0.14 a |
2.47±0.14 a |
16.55±0.99 a |
54.00±3.64 a |
|
60 |
1.42±0.17 a |
3.38±0.11 a |
3.33±0.16 a |
2.90±0.23 a |
19.86±1.09 b |
57.03±4.15 a |
|
120 |
1.69±0.19 a |
3.31±0.19 a |
3.56±0.18 a |
2.92±0.40 a |
20.06±1.04 b |
55.43±1.05 a |
|
180 |
2.10±0.15 a |
3.33±0.34 a |
3.70±0.62 a |
2.98±0.36 a |
19.87±1.83 b |
61.60±3.41 a |
|
HSD0.05 |
2.17 |
|||||
|
AMF Dose (g plant-1) |
||||||
|
0 |
1.64±0.49 a |
3.12±3.12 a |
3.19±0.09 a |
2.51±0.15 a |
18.21±2.20 a |
54.13±2.52 a |
|
10 |
1.57±0.27 a |
3.25±3.25 ab |
3.50±0.06 ab |
2.88±0.33 ab |
19.26±1.11 a |
56.21±4.30 ab |
|
20 |
1.80±0.27 a |
3.52±3.52 b |
3.69±0.55 b |
3.06±0.32 b |
19.78±2.00 a |
60.70±3.08 b |
|
HSD0.05 |
0.37 |
0.47 |
0.53 |
6.29 |
||
Description: Numbers followed by the same letter in the same column are not significantly different at the 5% significance level (HSD0.05) for each treatment.
3.1.2 Number of chlorophyll leaves, percentage of AMF colonization, weight of fresh biomass, root length, and root weight of sweet potato plants at age 6 WAP
At 6 WAP, chlorophyll content tended to be lower in plants receiving 180 kg ha-1 K₂O. AMF colonization, root length, and root weight did not differ significantly among treatments, likely because potassium application did not exceed 180 kg ha-1 (Table 3). This is consistent with Putra and Permadi [8], who reported that the effect of increasing K2O peaks at 150-225 kg ha-1, during which leaf photosynthetic rates initially rise before declining. Chlorophyll content is a reliable indicator of nitrogen uptake [28], as supported by Shu et al. [29], who found that SPAD values in sweet potato leaves increased with higher nitrogen application. The lack of a significant potassium effect on chlorophyll content may be attributed to soil chemical properties (pH), moisture status, and unfavorable environmental conditions, particularly low rainfall at 6 WAP (Table 1), which could have limited nutrient uptake.
Biomass fresh weight and root weight were highest in the 180 kg ha⁻¹ K₂O treatment, though not significantly different from the 120 kg ha⁻¹ treatment (Table 3). This finding is consistent with a previous report [30], which observed a linear increase in fresh biomass with increasing potassium doses. Potassium application enhanced vegetative organ development compared to the control, likely due to its role in promoting cell enlargement and elongation. In contrast, potassium deficiency can impair photosynthesis and reduce assimilate production, limiting the formation of plant organs such as leaves [31].
Table 3 also shows that chlorophyll content tended to be higher in the control (without AMF) but was not significantly different from other treatments. This contrasts with Lone et al. [19], who found higher chlorophyll content in AMF-inoculated plants. The relatively low chlorophyll values observed here may be due to measurements taken at the end of the vegetative stage (6 WAP), when chlorophyll levels may have already declined.
Table 3. Average chlorophyll number, percentage of AMF colonization, weight of fresh biomass, root length, and root weight of sweet potato plants at 6 WAP due to potassium fertilizer and AMF dosage
|
K2O Dose (kg ha-1) |
Number of Chlorophylls |
AMF Colonization (%) |
Weight of Fresh Biomass (g plant-1) |
Root Length (cm) |
Root Weight (g plant-1) |
|
0 |
49.04±0.65 a |
62.22±34.57 a |
198.23±17.71 a |
38.00±0.27 a |
5.41±0.50 a |
|
60 |
49.91±0.63 a |
55.56±39.38 a |
201.56±2.05 a |
43.33±7.76 a |
5.67±1.73 a |
|
120 |
49.94±1.06 a |
62.22±44.00 a |
251.77±4.32 ab |
45.89±6.56 a |
6.07±0.56 a |
|
180 |
48.43±0.97 a |
71.11±40.86 a |
252.27±24.85b |
46.17±1.77 a |
6.16±0.09 a |
|
HSD0.05 |
53.70 |
||||
|
AMF Dose (g plant-1) |
|||||
|
0 |
50.04±0.83 a |
6.67±6.67 a |
211.85±30.77 a |
39.25±5.50 a |
5.72±1.02 a |
|
10 |
48.81±1.28 a |
91.67±5.53 b |
230.52±25.44 a |
46.42±6.03 a |
5.36±0.51 a |
|
20 |
49.13±0.48 a |
90.00±7.45 b |
235.51±29.20 a |
44.38±4.25 a |
6.41±1.04 a |
|
HSD0.05 |
0.37 |
Description: Numbers followed by the same letter in the same column are not significantly different at the 5% significance level (HSD0.05) for each treatment.
At 6 WAP, AMF colonization and root length were significantly higher in plants receiving 10 g plant⁻¹ AMF than in the control, but not different from the 20 g plant⁻¹ treatment. Colonization exceeded 90% in both AMF treatments (91.67% at 10 g plant⁻¹ and slightly higher at 20 g plant⁻¹), indicating strong fungal infectivity. Fresh biomass and root weight tended to be greater with 20 g plant⁻¹ AMF, though differences from other treatments were not significant (Table 3). Similar to findings in potato cultivars [19], this suggests compatibility between local sweet potato clones and AMF. AMF plants generally develop longer roots with greater surface area and volume than non-AMF plants [32], a trend observed here and likely driven by enhanced phosphorus uptake in colonized roots [19].
3.2 Effect of potassium fertilizer dose and AMF on weight of fresh biomass, weight of dry biomass, number of tubers, tuber length, and tuber diameter of sweet potato plants
3.2.1 Weight of fresh biomass, and weight of dry biomass
Table 4 shows that the highest fresh and dry biomass of sweet potato plants was obtained at a potassium dose of 180 kg ha⁻¹, which was significantly different from the control. Increased N and K accumulation significantly enhanced plant growth, while greater potassium availability also promoted higher dry matter accumulation in shoots and roots. This increase in whole-plant dry matter serves as the foundation for achieving higher sweet potato yields [10].
Table 4 also indicates that the fresh and dry biomass of sweet potato plants was higher with an AMF dose of 20 g plant⁻¹, which was not significantly different from the 10 g plant⁻¹ treatment but was significantly higher than the control. AMF promote root growth and the development of denser, more highly branched root systems by increasing root surface area, thereby enhancing the plant’s ability to absorb water and nutrients from the soil [33], which in turn supports overall plant growth. It has been reported that AMF symbiosis can improve the uptake of both macro- and micronutrients, particularly those present in bound or less available forms [34].
Table 4. Average weight of fresh biomass, weight of dry biomass, number of tubers, tuber length, and tuber diameter of sweet potato plants at 17 WAP due to potassium fertilizer and AMF dosage
|
K2O Dose (kg ha-1) |
Weight of Fresh Biomass (g plant-1) |
Weight of Dry Biomass (g plant-1) |
Number of Tubers (Tuber plant-1) |
Tuber Length (cm) |
Tuber Diameter (mm) |
|
0 |
380.82±103.99 a |
75.45±8.10 a |
4.58±0.11 a |
13.81±0.81 a |
53.06±7.64 a |
|
60 |
459.16±99.49 ab |
75.46±3.16 a |
4.67±0.24 a |
14.48±0.64 ab |
48.67±1.63 a |
|
120 |
460.34±29.08 ab |
77.43±3.16 ab |
4.96±0.17 ab |
15.00±0.36 abc |
57.18±2.23 a |
|
180 |
514.66±18.68 b |
90.20±9.27 b |
5.64±0.40 b |
16.12±0.83 c |
56.53±0.60 a |
|
HSD0.05 |
107.41 |
14.11 |
0.66 |
1.38 |
|
|
AMF Dose (g plant-1) |
|||||
|
0 |
397.65±73.07 a |
73.77±3.12 a |
4.88±0.26 a |
14.42±0.44 a |
53.54±4.38 a |
|
10 |
423.55±71.25 ab |
78.38±8.86 ab |
4.88±0.36 a |
14.57±1.41 ab |
51.78±3.91 a |
|
20 |
540.04±36.53 b |
86.76±8.19 b |
5.12±0.70 a |
15.57±0.77 b |
56.26±6.27 a |
|
HSD0.05 |
83.39 |
11.73 |
1.07 |
Description: Numbers followed by the same letter in the same column are not significantly different at the 5% significance level (HSD0.05) for each treatment.
3.2.2 Number of tubers, tuber length, and tuber diameter
Table 4 shows that the number of tubers and tuber length were the highest at a potassium fertilizer dose of 180 kg ha-1, which was significantly different from the other treatments. Potassium affects tuber yield by increasing the proportion of photosynthetic dry matter transported to tubers. Based on yield components, increasing the dose of potassium fertilizer increased plant yield as indicated by the number of tubers, tuber length, and tuber weight. Study [35] stated that the most significant function of potassium in regulating the opening of stomata is to balance the entry of CO2 and remove H2O from the intercellular space. The amount of CO₂ entering the intercellular spaces reflects the level of photosynthate production in the leaf. Potassium is essential for activating the enzyme ATP synthase, thereby facilitating the photosynthetic process. According to study [35], ATPase activity reaches its optimum performance when the potassium content in the plant is at an optimal level.
The diameter of the tubers tended to be larger at the potassium fertilizer dose of 120 kg ha-1 (Table 4); however, this difference was not statistically significant compared to the other treatments. A study [36] found that applying 120 kg ha-1 significantly increased the diameter of tubers to 5.69 cm. The increase in yield due to K fertilization was mainly due to the increase in the tuber-to-plant ratio, leading to more translocation of photosynthate to the tuber and increasing its size.
Table 4 shows that the length of sweet potato tubers was higher in the AMF of 20 g plant-1 and was not significantly different from 10 g plant-1 but significantly different from the control. Similarly, study [37] found that plants treated with AMF can form longer tubers compared to other treatments. The number of tubers and tuber diameter tended to be higher in the AMF of 20 g plant-1, but not significantly different from the other treatments. Study [25] reported that AMF can significantly enhance stem length, leaf number, branch number, as well as tuber weight, diameter, and length. AMF improve phosphorus availability by breaking down soil-bound P, facilitated by hyphal secretion of phosphatase enzymes that release P into forms accessible to roots. This enhanced P uptake not only promotes greater branch development in sweet potato but also allows AMF-infected plants to absorb more phosphorus compared to non-infected plants [38].
3.3 Effect of potassium fertilizer dose and AMF on tuber weight, yield, harvest index, starch, and β-carotene content of sweet potato plants
3.3.1 Tuber weight and yield
Tuber weight and yield of sweet potato plants were higher at the potassium dose of 180 kg ha-1. It was not significantly different from the dose of 120 kg ha-1, but it was significantly different from the control and the potassium dose of 60 kg ha-1 (Table 5). This indicates that the tuber weight has not reached the optimum value at a dose of 180 kg ha-1, study [8] found that the tuber weight increased with the increase of K2O rate at harvesting, with the highest in the treatment of 225-300 kg ha-1. This increase is preceded by the supply of potassium, which can significantly increase the transport of N from the shoots to roots and increase the root N distribution ratio during the late period of tuber growth. The supply of potassium also decreases the N/K ratio of sweet potato and increases the transport of dry matter to the tuber in the final stage of the growth period [39]. The results of this study align with study [39], which reported optimum sweet potato yield at the potassium dose of 150-225 kg ha-1. Potassium affects tuber yield by increasing the allocation of dry matter to tubers and increasing the number, length, and diameter of tuber plants.
Potassium plays a role in almost all plant physiological processes, including water absorption, nitrogen use efficiency, stomatal regulation, enzyme activation, assimilate translocation, photosynthesis, and drought and disease resistance [40]. In addition, potassium aids water transport and the translocation of mineral compounds to the whole plant through the xylem. In cases where K supply is not at optimal levels, translocation of mineral compounds, such as nitrate (NO3-), phosphate (PO43-), calcium (Ca2+), magnesium (Mg2+), and amino acid uptake are reduced.
Table 5. Average tuber weight, yield, harvest Index, starch, and β-carotene content of sweet potato plants at 17 WAP due to potassium fertilizer and AMF doses
|
K2O Dose (kg ha-1) |
Tuber Weight (g plant-1) |
Yield (ton ha-1) |
Harvest Index |
Starch (%) |
β-Carotene (%) |
|
0 |
1032.76±301.81 a |
6.45±1.89 a |
0.730±0.007 a |
67.70±1.66 a |
2.83±0.13 b |
|
60 |
1096.79±68.45 ab |
6.85±0.43 ab |
0.704±0.054 a |
67.48±1.42 a |
1.75±0.13 a |
|
120 |
1342.50±143.43 bc |
8.39±0.90 bc |
0.744±0.021 a |
66.42±1.34 a |
3.32±0.16 c |
|
180 |
1483.04±202.97 c |
9.27±1.27 c |
0.741±0.018 a |
66.46±1.00 a |
2.69±0.12 b |
|
HSD0.05 |
305.52 |
1.91 |
0.21 |
||
|
AMF Dose (g plant-1) |
|||||
|
0 |
1127.21±225.55 a |
7.05±1.41 a |
0.739±0.018 a |
66.36±1.81 a |
2.73±0.54 a |
|
10 |
1163.67±231.92 ab |
7.27±1.45 ab |
0.732±0.014 a |
66.70±1.16 ab |
2.60±0.68 a |
|
20 |
1425.43±245.65 c |
8.91±1.54 c |
0.718±0.053 a |
67.98±0.76 b |
2.62±0.52 a |
|
HSD0.05 |
237.19 |
1.48 |
1.37 |
Description: Numbers followed by the same letter in the same column are not significantly different at 5% significance level (HSD0.05) for each treatment.
Table 5 shows that the tuber weight and yield of the sweet potato plant were higher in the treatment with an AMF 20 g plant-1, and were significantly different from other treatments. Various studies show the critical role of AMF in helping the availability and absorption of phosphorus in plants. With the process of tuber filling, higher tuber weights were obtained in AMF-inoculated plants. Given that tuber initiation is hormonally mediated [41], it is possible that in this study, AMF affected the hormonal balance in sweet potato.
The low yield, recorded below 10 tons ha-1, is likely due to suboptimal climatic conditions. Although the vegetative phase received adequate rainfall, the generative phase, particularly during tuber filling, coincided with the dry season and experienced low rainfall (Table 1). As shown in Table 1, during the vegetative period (April-May), the average of rainfall was 41.0 mm, the average of air temperature was 27.5℃ (maximum and minimum air temperature 34.9℃ and 22.6℃, respectively), and the average of air humidity was 80.2%. In contrast, the tuber formation period (June-September) occurred during the dry season, with the average of rainfall being 29.0 mm, the average air temperature was 27.7℃ (maximum and minimum air temperature 35.3℃ and 22.6℃, respectively), and the average of air humidity was 75.8%.
The maximum number of tubers increased quadratically with rising temperature, peaking at 25.3℃, and decreased with further temperature increases. A similar trend was observed for the percentage of tubers relative to the total number of adventitious roots produced, which reached its maximum at 23.9℃ [42]. Sweet potato thrives in warm tropical climates with an average temperature of around 24℃ [1] and well-distributed annual rainfall between 1000 and 2000 mm. Climatic conditions characterized by high temperatures and limited water availability during the dry season can disrupt plant metabolic processes. Photosynthetic efficiency largely depends on sufficient water and nutrient supply. Under water stress, reduced stomatal conductance hampers photosynthesis and may result in substantial yield losses [43].
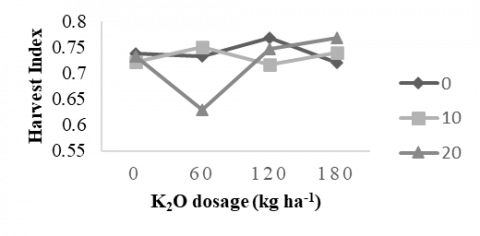
3.3.2 Harvest index
Figure 1 shows the average harvest index (HI) of sweet potato due to the interaction between potassium dose and AMF treatment harvested at 17 WAP. The overall average HI in this study was 0.73. The best combination between potassium dose and AMF on sweet potato HI was found in a potassium dose of 120 kg ha-1 without AMF (0.77). However, it was not significantly different from the treatment combination of a potassium dose of 60 kg ha-1 with or without AMF (0.73). At a K2O fertilization dose of 120 kg ha-1, the harvest index was 0.74, and with an AMF dose of 10 g plant-1, it was 0.73.
The harvest index (HI) reflects a plant’s efficiency in converting photosynthates into marketable products. In sweet potato, the tuber is the primary commercial product for food and industrial use, although branches and leaves also have value as animal feed. Tubers rich in starch and sugar are economically important, but their HI is challenging to determine due to factors such as their underground location and the influence of harvest timing. Delayed harvesting beyond physiological maturity can result in losses of aboveground dry matter through aging or senescence. HI is a key parameter for assessing yield potential and cultivation efficiency, with higher values indicating more effective allocation of assimilates to harvestable organs. It represents the proportion of total biomass partitioned to the tuber relative to aboveground biomass. Across studies, HI in 15 sweet potato clones ranged from 0.51 to 0.88 [44], with superior varieties generally exhibiting higher values than local clones. Similarly, research in Nigeria reported HI values up to 0.53 in improved varieties, exceeding the grand mean of 0.46 for local landraces [45].
Figure 1. Interaction between potassium dose and AMF at 17 WAP on the harvest index value in sweet potato plants
In this study, harvest index values ranged from 0.63 to 0.77. These values are comparable to those reported by [46], who found a sweet potato HI of 0.6 due to NPK fertilization and agro-ecological variations studied in Ghana. They recommended this rate as the optimal potassium dose due to its significant economic benefits and theoretical value. [20] studied the effect of K fertilizer on sweet potato. They found that, compared to other treatments (100 and 300 kg ha-1 K2O), the 200 kg ha-1 application resulted in higher yield, starch content, and net profit of sweet potato.
The weight of fresh tubers, measured in fresh form, includes water content and plant parts that undergo varying degrees of water loss. Study [45] evaluated nine local sweet potato accessions of Nigerian origin for potential selection as varieties based on HI and tuber weight. Aboveground biomass influences HI as it is responsible for transporting the photosynthate allocated to storage organs (sink/tubers) and to the aboveground plant parts. The proportion of photosynthate allocated to the sink is genetically determined, resulting in variation in the HI among sweet potato genotypes.
The research results of study [47] concluded that potassium and AMF (Claroideoglomus etunicatum) showed synergistic effects on tuber development and potassium acquisition at high potassium use efficiency. The use of AMF will increase the HI with potassium use. Study [48] reported that the enhancement of tuber production is closely linked to the role of AMF in improving nutrient absorption, stimulating photosynthate production, and increasing biomass accumulation. The rise in photosynthetic activity observed in AMF-inoculated plants is directly associated with greater uptake of nitrogen (N), phosphorus (P), and carbon, which collectively contribute to improved tuber development.
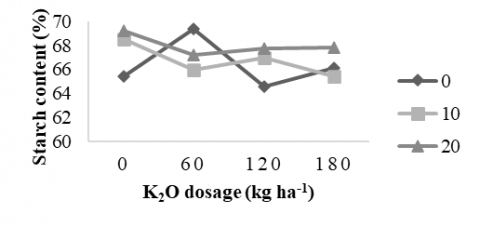
3.3.3 Starch content
The average starch content of sweet potato, resulting from the positive interaction between potassium dosage and AMF treatment at 17 WAP, is presented in Figure 2. Figure 2 shows that the best treatment combination between potassium dose and AMF on sweet potato starch content was found in a potassium dose of 60 kg ha-1 without AMF (69.36%), but not significantly different from the AMF dose of 20 g plant-1 without potassium (69.22%). Meanwhile, in individual treatment at a K2O fertilization dose of 120 kg ha-1, the starch content was 66.42% and with an AMF dose of 10 g plant-1, it was 66.70% (Table 5). This result is consistent with the findings of study [8], which reported that potassium application reduced the amylose and amylopectin content in tubers compared with the control.
Specifically, K application decreased amylose and amylopectin content by 9–13% and 6–11% in Xushu 32, respectively, and by 10–23% and 13–19% in Ningzishu 1. However, potassium can also improve the taste, shape, size, color, and texture of tubers [49]. Starch, the primary carbohydrate stored in tubers and root crops, is synthesized from sucrose degradation [50]. This conversion is regulated by several key enzymes, with sucrose phosphatase playing a central role in sucrose biosynthesis by controlling the allocation and conversion of photosynthates between sucrose and starch. Meanwhile, invertase and sucrose synthase are essential for sucrose degradation, enabling its utilization in starch polymer formation. Together, these metabolic enzymes play a central role in the sucrose-to-starch conversion process, with their encoding genes being strongly associated with starch accumulation [51].
Figure 2. Interaction between potassium dose and AMF at 17 WAP on starch content (%) in sweet potato plants
The results in Figure 2 indicate that higher doses of AMF lead to increased starch content. This effect may be attributed to the enhanced translocation of assimilates to roots and tubers, which function as major sinks. Similar trends have been reported, with AMF application resulting in higher starch accumulation in potatoes [19], and further increases observed when AMF was combined with potassium doses of 120 kg ha⁻¹ and 180 kg ha⁻¹. AMF symbiosis enhances sucrose transport, as evidenced by decreased sucrose content in shoots and increased sucrose content in roots. Through its role in regulating the source–sink relationship, AMF facilitates the transfer of carbohydrates from source tissues to tubers, resulting in greater tuber weight compared to the control.
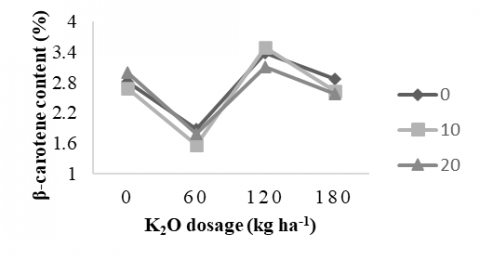
3.3.4 β-carotene content
Figure 3 illustrates the average β-carotene content of sweet potatoes affected by the interaction of potassium fertilization and AMF treatment at 17 WAP. The potassium dose of 120 kg ha⁻¹ without AMF resulted in the optimal combination for enhancing β-carotene content in sweet potato (3.37%). With K2O fertilization at a dose of 120 kg ha-1 alone, or an AMF dose of 10 g plant-1 alone, the β-carotene levels were 3.32% and 2.60% respectively (Table 5). These results indicate that adequate potassium supply increases carotenoid biosynthesis, especially β-carotene, by stimulating photosynthetic efficiency and assimilate translocation to storage roots. However, the addition of AMF under high potassium conditions did not provide a synergistic effect, possibly due to the reduced dependence of plants on AMF in nutrient-rich environments. This is by limiting the total root length colonized by AMF [52]. The absence of a significant synergistic effect between potassium fertilization and AMF inoculation is likely influenced by initial soil nutrient conditions, particularly phosphorus availability, which can suppress AMF activity. Under drought stress, reduced root growth and microbial activity further constrain AMF colonization and potassium mobility. These factors together may explain the lack of synergy, highlighting the critical role of soil fertility and environmental stress in shaping plant responses to nutrient and microbial inputs.
Figure 3. Interaction between potassium dose and AMF at 17 WAP on β-carotene content (%) in sweet potato plants
Potassium at a dose of 120 kg ha-1 increases tuber yield and enhances β-carotene content. This finding is supported by study [6], which reported an increase in both β-carotene content and tuber yield, reaching up to 10.55 ton ha-¹ in the 'Narutokintoki' variety cultivated in paddy fields. However, study [53] recommended a lower dose of 73-77 kg ha-1 K2O to optimize total carotenoids, β-carotene, and α-carotene in the 'Beauregard' cultivar. Specifically, potassium affects enzymes involved in the β-carotene biosynthesis pathway, helping plants produce more of this pigment.
The findings indicate that applying 120 kg ha⁻¹ K₂O is an optimal strategy for maximizing yield and enhancing nutritional quality, particularly β-carotene content in local sweet potato genotypes, providing an evidence-based guideline for fertilizer management in similar agroecological zones; however, while AMF inoculation improved growth, its inconsistent effects on quality under low-rainfall conditions highlight the need to evaluate its performance across different seasons and irrigation regimes and underscore the importance of aligning nutrient inputs and planting schedules with prevailing climatic patterns to optimize resource use efficiency. Further research suggests conducting experiments during the rainy season to investigate the interactive effect under well-watered conditions.
The authors gratefully acknowledge Universitas Syiah Kuala for financial support provided through Research Grant No. 202/UN11.2.1/PT.01.03/PNBP/2023. Special thanks are extended to the field team, Arfan Ependi and Saparuddin, for their invaluable assistance with field activities, including planting, maintenance, harvesting, and data collection.
[1] Sapakhova, Z., Raissova, N., Daurov, D., Zhapar, K., Daurova, A., Zhigailov, A., Zhambakin, K., Shamekova, M. (2023). Sweet potato as a key crop for food security under the conditions of global climate change: A review. Plants, 12(13): 2516. https://doi.org/10.3390/plants12132516
[2] Laveriano-Santos, E.P., López-Yerena, A., Jaime-Rodríguez, C., González-Coria, J., Lamuela-Raventós, R.M., Vallverdú-Queralt, A., Romanyà, J., Pérez, M. (2022). Sweet potato is not simply an abundant food frop: A comprehensive review of its phytochemical constituents, biological activities, and the effects of processing. Antioxidants, 11(9): 1648-1675. https://doi.org/10.3390/antiox11091648
[3] BPS. (2024). Indonesian Statistic 2023. Indonesian Central Bureau of Statistics. Jakarta. https://www.bps.go.id/id.
[4] Hayati, M., Sabaruddin, Efendi, Anhar, A. (2019). Chemical and sensory analysis of several sweet potatoes (Ipomoea batatas L.) clones. IOP Conference Series: Earth and Environmental Science, 364(1): 012011. https://doi.org/10.1088/1755-1315/364/1/012011
[5] Hayati, M., Sabaruddin, Efendi, Anhar, A. (2020). Morphological characteristics and yields of several sweet potato (Ipomoea batatas L.) tubers. IOP Conference Series: Earth and Environmental Science, 425(1): 012055. https://doi.org/10.1088/1755-1315/425/1/012055
[6] Tang, Z.H., Zhang, A.J., Wei, M., Chen, X.G., Liu, Z.H., Li, H.M., Ding, Y.F. (2015). Physiological response to potassium deficiency in three sweet potato (Ipomoea batatas [L.] Lam.) genotypes differing in potassium utilization efficiency. Acta Physiologiae Plantarum, 37(8): 184. https://doi.org/10.1007/s11738-015-1901-0
[7] Agung, M., Sulistyawati, S., Purnamasari, R.T. (2020). The effect of different varieties and doses of potassium fertilizer on the growth and production of sweet potato plants (Ipomoea batatas L.). Jurnal Agroteknologi Merdeka Pasuruan, 4(1): 8-13. https://jamp-jurnal.unmerpas.ac.id/index.php/jamppertanian/article/view/38.
[8] Putra, S., Permadi, K. (2011). The effect of potassium fertilizer on increasing the yield of sweet potato variety Narutokintokidi in rice fields. Agrin, 15(2): 133-142. https://jurnalagrin.net/index.php/agrin/article/view/189/158.
[9] Gao, Y., Tang, Z., Xia, H., Sheng, M., Liu, M., Pan, S., Li, Z., Liu, J. (2021). Potassium fertilization stimulates sucrose-to-starch conversion and root formation in sweet potato (Ipomoea batatas (L.) Lam.). International Journal of Molecular Sciences, 22(9): 4826. https://doi.org/10.3390/ijms22094826
[10] El-Baky, A., Ahmed, A.A., El-Nemr, M.A., Zaki, M.F. (2010). Effect of potassium fertilizer and foliar zinc application on yield and quality of sweet potato. Research Journal of Agriculture and Biological Sciences, 6(4): 386-394.
[11] Alaboz, P., Dengiz, O., Pacci, S., Demir, S., Türkay, C. (2022). Determination of the effect of different organic fertilizers applications on soil quality using the SMAF model. Yuzuncu Yil University Journal of Agricultural Sciences, 32(1): 21-32. https://doi.org/10.29133/yyutbd.1015943
[12] Bunn, R.A., Simpson, D.T., Bullington, L.S., Lekberg, Y., Janos, D.P. (2019). Revisiting the ‘direct mineral cycling’ hypothesis: Arbuscular mycorrhizal fungi colonize leaf litter, but why? ISME Journal, 13: 1891-1898. https://doi.org/10.1038/s41396-019-0403-2
[13] Tong, Y., Gabriel-Neumann, E., Ngwene, B., Krumbein, A., Baldermann, S., Schreiner, M., George, E. (2013). Effects of single and mixed inoculation with two arbuscular mycorrhizal fungi in two different levels of phosphorus supply on β-carotene concentrations in sweet potato (Ipomoea batatas L.) tubers. Plant and Soil, 372(1): 361-374. https://doi.org/10.1007/s11104-013-1708-y
[14] de Albuquerque, J.R.T., Ribeiro, R.M.P., Pereira, L.A.F., Junior, A.P.B., da Silveira, L.M., dos Santos, M.G., de Souza, A.R.E., Lins, H.A., Neto, F.B. (2016). Sweet potato cultivars grown and harvested at different times in semiarid Brazil. African Journal of Agricultural Research, 11(46): 4810-4818. https://doi.org/10.5897/ajar2016.11761
[15] Othman, R., Kammona, S., Jaswir, I., Jamal, P., Mohd Hatta, F.A. (2017). Influence of growing location, harvesting season and post-harvest storage time on carotenoid biosynthesis in orange sweet potato (Ipomoea batatas) tuber flesh. International Food Research Journal, 24: 488-495.
[16] Singogo, B., Mataa, M., Munyinda, K., Lungu, D., Dairy, P., Box, P.O. (2021). Variation in beta carotene and yield in sweetpotato (Ipomoea batatas (L.) Lam.) associated with ambient temperature and genotype. African Journal of Agricultural Research, 17(3): 479-486. https://doi.org/10.5897/AJAR2020.14927
[17] Davar, R., Sabbaghtazeh, E., Bybordi, A., Dalalian, M.R., Saedi, S. (2023). Application of arbuscular mycorrhizal fungi and potassium nitrate improves physiological performance and glycyrrhizin production of licorice under salt stress. Journal of Applied Botany and Food Quality, 96: 130-137. https://doi.org/10.5073/JABFQ.2023.096.017
[18] Sakha, M.A., Jefwa, J., Gweyi-Onyango, J.P. (2019). Effects of arbuscular mycorrhizal fungal inoculation on growth and yield of two sweet potato varieties. Journal of Agriculture and Ecology Research International, 18(3): 1-8. https://doi.org/10.9734/jaeri/2019/v18i330063
[19] Lone, R., Shuab, R., Sharma, V., Kumar, V., Mir, R., Koul, K.K. (2015). Effect of arbuscular mycorrhizal fungi on growth and development of potato (Solanum tuberosum) plant. Asian Journal of Crop Science, 7(3): 233-243. https://doi.org/10.3923/ajcs.2015.233.243
[20] Geng, J., Zhao, Q., Li, Z., Yang, X., Lei, S., Zhang, Q., Li, H., Lang, Y., Huo, X., Liu, Q. (2024). Effects of various potassium fertilizer dosages on agronomic and economic assessment of sweet potato fields. Horticulturae, 10(1): 44-55. https://doi.org/10.3390/horticulturae10010044
[21] Uwah, D.F., Undie, U.L., John, N.M., Ukoha, G.O. (2013). Growth and yield response of improved sweet potato (Ipomoea batatas (L.) Lam) varieties to different rates of potassium fertilizer in Calabar, Nigeria. Journal of Agricultural Science, 5(7): 61-69. https://doi.org/10.5539/jas.v5n7p61
[22] Abd El-Latif, K.M., Osman, E.A.M., Abdullah, R., Abd El-Kader, N. (2011). Response of potato plants to potassium fertilizer rates and soil moisture deficit. Advances in Applied Science Research, 2(2): 388-397. https://doi.org/10.21608/jpp.2018.35254
[23] Trehan, S.P., Pandey, S.K., Bansal, S.K. (2009). Potassium nutrition of potato crops-the Indian Scenario. Optimizing Crop Nutrition, 19: 2-9. https://www.ipipotash.org/publications/eifc-11.
[24] Xu, X., Du, X., Wang, F., Sha, J., Chen, Q., Tian, G., Zhu, Z., Ge, S., Jiang, Y. (2020). Effects of potassium levels on plant growth, accumulation and distribution of carbon, and nitrate metabolism in apple dwarf rootstock seedlings. Frontiers in Plant Science, 11: 904. https://doi.org/10.3389/fpls.2020.00904
[25] Pelealu, J.J., Wahyudi, L., Tallei, T.E. (2019). Growth response and production of purple sweet potatoes after provision of arbuscular mycorrhizal fungi and organic fertilizer. Asian Journal of Plant Sciences, 18(3): 123-130. https://doi.org/10.3923/ajps.2019.123.130
[26] Whiteside, M.D., Digman, M.A., Gratton, E., Treseder, K.K. (2012). Organic nitrogen uptake by arbuscular mycorrhizal fungi in a boreal forest. Soil Biology and Biochemistry, 55: 7-13. https://doi.org/10.1016/j.soilbio.2012.06.001
[27] Wahab, A., Muhammad, M., Munir, A., Abdi, G., Zaman, W., Ayaz, A., Khizar, C., Reddy, S.P.P. (2023). Role of arbuscular mycorrhizal fungi in regulating growth, enhancing productivity, and potentially influencing ecosystems under abiotic and biotic Stresses. Plants, 12(17): 3102-3141. https://doi.org/10.3390/plants12173102
[28] Yeboah, S., Zhang, R., Cai, L., Li, L., Xie, J., Luo, Z., Wu, J., Antille, D.L. (2017). Soil water content and photosynthetic capacity of spring wheat as affected by soil application of nitrogen-enriched biochar in a semiarid environment. Photosynthetica, 55(3): 532-542. https://doi.org/10.1007/s11099-016-0672-1
[29] Shu, X., Jin, M., Wang, S., Xu, X., Deng, L., Zhang, Z., Zhao, X., Yu, J., Zhu, Y., Lu, G., Lv, Z. (2024). The effect of nitrogen and potassium interaction on the leaf physiological characteristics, yield, and quality of sweet potato. Agronomy, 14(10): 2319-2333. https://doi.org/10.3390/agronomy14102319
[30] Suwarto, Aswidinoor, H., Reyzaldi, F.G. (2022). Effect of potassium fertilizers doses on the growth, tuber yield, and resistance to weevil infestation of sweet potato on Latosol soil. Australian Journal of Crop Science, 16(1): 137-142. https://doi.org/10.21475/ajcs.22.16.01.p3375
[31] Ning, C.C., Gao, P.D., Wang, B.Q., Lin, W.P., Jiang, N.H., Cai, K.Z. (2017). Impacts of chemical fertilizer reduction and organic amendments supplementation on soil nutrient, enzyme activity and heavy metal content. Journal of Integrative Agriculture, 16(8): 1819-1831. https://doi.org/10.1016/S2095-3119(16)61476-4
[32] Wu, Q.S., Li, G.H., Zou, Y.N. (2011). Roles of arbuscular mycorrhizal fungi on growth and nutrient acquisition of peach (Prunus persica L. Batsch) seedlings. The Journal of Animal & Plant Sciences, 21(4): 746-750.
[33] Ramírez-Flores, M.R., Bello-Bello, E., Rellán-Álvarez, R., Sawers, R.J., Olalde-Portugal, V. (2019). Inoculation with the mycorrhizal fungus Rhizophagus irregularis modulates the relationship between root growth and nutrient content in maize (Zea mays ssp. mays L.). Plant Direct, 3(12): e00192. https://doi.org/10.1002/pld3.192
[34] Liu, H., Shi, C.Y., Zhang, H., Wang, Z.Z., Chai, S. (2013). Effects of potassium on yield, photosynthate distribution, enzymes’ activity and ABA content in storage roots of sweet potato (Ipomoea batatas Lam.). Australian Journal of Crop Science, 7(6): 735-743.
[35] Hasanuzzaman, M., Bhuyan, M.H.M.B., Nahar, K., Hossain, M.S., Al Mahmud, J., Hossen, M.S., Masud, A.A.C., Moumita, Fujita, M. (2018). Potassium: A vital regulator of plant responses and tolerance to abiotic stresses. Agronomy, 8(3): 31. https://doi.org/10.3390/agronomy8030031
[36] Singh, P., Aravindakshan, K., Maurya, I.B., Singh, J., Singh, B., Sharma, M.K. (2017). Effect of potassium and zinc on growth, yield and economics of sweet potato (Ipomoea batatas L.) cv. CO-34. Journal of Applied and Natural Science, 9(1): 291-297. https://doi.org/10.31018/jans.v9i1.1186
[37] Rahmawati, N., Sitepu, F.E., Tarigan, M.F.A. (2021). Effectiveness of mycorrhizae on growth and production of orange sweet potato at various watering level. IOP Conference Series: Earth and Environmental Science, 782(4): 042046. https://doi.org/10.1088/1755-1315/782/4/042046
[38] Al Hadidi, N., Pap, Z., Ladányi, M., Szentpéteri, V., Kappel, N. (2021). Mycorrhizal inoculation effect on sweet potato (Ipomoea batatas (L.) Lam) seedlings. Agronomy, 11(10): 2019-2028. https://doi.org/10.3390/agronomy11102019
[39] Wang, S., Li, H., Liu, Q., Hu, S., Shi, Y. (2019). Nitrogen uptake, growth and yield response of orange-fleshed sweet potato (Ipomoea batatas L.) to potassium supply. Communications in Soil Science and Plant Analysis, 51(2): 175-185. https://doi.org/10.1080/00103624.2019.1695821
[40] Pushpalatha, M., Vaidya, P.H., Sunil, B.H., Adsul, P.B. (2018). Effect of graded levels of nitrogen and potassium on growth, nutrient content and uptake of sweet potato (Ipomoea batatas L.) in vertisols of maharashtra. International Journal of Plant & Soil Science, 23(5): 1-8. https://doi.org/10.9734/ijpss/2018/42408
[41] Parvizi, K., Dashti, F. (2015). The effect of in vitro mycorrhization on growth characteristics, changes in endogenous hormones and performance of microplants in potato (Solanum tuberosum L.). Journal of Central European Agriculture, 16(4): 445-462. https://doi.org/10.5513/JCEA01/16.4.1652
[42] Gajanayake, B., Reddy, K.R., Shankle, M.W., Arancibia, R.A., Villordon, A.O. (2014). Quantifying storage root initiation, growth, and developmental responses of sweetpotato to early season temperature. Agronomy Journal, 106(5): 1795-1804. https://doi.org/10.2134/agronj14.0067
[43] Van Heerden, P.D.R., Laurie, R. (2008). Effects of prolonged restriction in water supply on photosynthesis, shoot development and storage root yield in sweet potato. Physiologia Plantarum, 134(1): 99-109. https://doi.org/10.1111/j.1399-3054.2008.01111.x
[44] Rahajeng, W., Restuono, J., Indriani, F.C., Purwono, P. (2021). Evaluation of promising sweet potato clones for higher root yield and dry matter content. Journal of Agro Science, 9(1): 42-47. https://doi.org/10.18196/pt.v9i1.6026
[45] Nwankwo, I.I.M., Akinbo, O., Ikoro, A.I., Orji, N.A.C., Njoku, T. (2018). Evaluation of selected sweet potato landraces for high harvest index and high root yield indices for parental selection. International Journal of Agricultural Policy and Research, 6(7): 90-97. https://doi.org/10.15739/IJAPR.18.011
[46] Darko, C., Yeboah, S., Amoah, A., Opoku, A., Berchie, J.N. (2020). Productivity of sweet potato (Ipomoea batatas (L) Lam) as influenced by fertilizer application in different agro-ecologies in Ghana. Scientific African, 10: e00560. https://doi.org/10.1016/j.sciaf.2020.e00560
[47] Yuan, J., Shi, K., Zhou, X., Wang, L., Xu, C., Zhang, H., Zhu, G., Si, C., Wang, J., Zhang, Y. (2023). Interactive impact of potassium and arbuscular mycorrhizal fungi on the root morphology and nutrient uptake of sweet potato (Ipomoea batatas L.). Frontiers in Microbiology, 13: 1075957. https://doi.org/10.3389/fmicb.2022.1075957
[48] Begum, N., Qin, C., Ahanger, M.A., Raza, S., Khan, M.I., Ashraf, M., Ahmed, N., Zhang, L. (2019). Role of arbuscular mycorrhizal fungi in plant growth regulation: implications in abiotic stress tolerance. Frontiers in Plant Science, 10: 1068. https://doi.org/10.3389/fpls.2019.01068
[49] Nedunchezhiyan, M., Byju, G., Ray, R.C. (2012). Effect of tillage, irrigation, and nutrient levels on growth and yield of sweet potato in rice fallow. International Scholarly Research Notices. ISRN Agronomy, 2012(1): 291285. https://doi.org/10.5402/2012/291285
[50] Liu, J., Ma, Y., Lv, F., Chen, J., Zhou, Z., Wang, Y., Abudurezike, A., Oosterhuis, D.M. (2013). Changes of sucrose metabolism in leaf subtending to cotton boll under cool temperature due to late planting. Field Crops Research, 144: 200-211. https://doi.org/10.1016/j.fcr.2013.02.003
[51] Kannan, C.G., Perumalsamy, P., Thangavelu, M. (2017). Influences of potassium chloride fertilization on mycorrhizal formation in a tropical alfisol. Communications in Soil Science and Plant Analysis, 48(5): 524-538. https://doi.org/10.1080/00103624.2016.1269789
[52] Sun, T., Rao, S., Zhou, X., Li, L. (2022). Plant carotenoids: recent advances and future perspectives. Molecular Horticulture, 2(1): 1-21. https://doi.org/10.1186/s43897-022-00023-2
[53] Nascimento, S.M.C., Cecílio-Filho, A.B., Silva, A.S.N.D., Vargas, P.F. (2019). Carotene yield in sweet potato after potassium and phosphorus fertiliser application. Revista Caatinga, 32(3): 851-857. https://doi.org/10.1590/1983-21252019v32n330rc