Ana Amiroh![]() | Edi Purwanto
| Edi Purwanto![]() | Eddy Tri Haryanto
| Eddy Tri Haryanto![]() | Ahmad Yunus*
| Ahmad Yunus*![]()
© 2025 The authors. This article is published by IIETA and is licensed under the CC BY 4.0 license (http://creativecommons.org/licenses/by/4.0/).
OPEN ACCESS
Watermelon (Citrullus lanatus L.) is highly susceptible to drought stress, which adversely affects its physiological and biochemical performance. This study aimed to assess the effectiveness of organic ameliorants—water hyacinth bokashi, manure, and rice husk biochar—in mitigating the impacts of drought stress on watermelon. A factorial randomized block design (RBD) was employed with two factors: (1) four combinations of organic ameliorants—BoM (bokashi + manure), BoBi (bokashi + biochar), MBi (manure + biochar), and BoMBi (bokashi + manure + biochar); and (2) four levels of drought stress based on field capacity (FC)—100%, 75%, 50%, and 25%. Results demonstrated that certain ameliorant combinations, particularly MBi and BoM under 25% FC (MBi25 and BoM25), significantly sustained higher levels of chlorophyll a (127.57 µg/g and 135.84 µg/g, respectively; p < 0.05), chlorophyll b (42.40 µg/g and 48.29 µg/g; p < 0.05), and total chlorophyll (169.97 µg/g and 184.13 µg/g; p < 0.05), indicating improved photosynthetic capacity under severe drought conditions. Additionally, BoM25 maintained a relatively high leaf water content (69.70%), despite root growth inhibition. In contrast, proline, antioxidant activity, and total phenolic content were markedly elevated under extreme drought (25% FC), although the ameliorants had limited impact on these stress markers. Overall, the application of organic ameliorants partially alleviated the negative effects of drought stress by enhancing water retention and preserving chlorophyll content, while biochemical stress indicators remained largely unaffected.
abiotic stress, drought tolerance, soil amendments, Citrullus lanatus, physiological responses, organic
Drought is a major constraint to agricultural productivity worldwide and is regarded as one of the most severe and widespread abiotic stresses affecting crop growth and yield [1-3]. Climate change has exacerbated the frequency and intensity of drought events, posing a significant threat to food security, particularly in arid and semi-arid regions. According to projections, global freshwater resources are expected to decline by up to 30% by the year 2050, and the total land area affected by drought may double [4]. This pressing issue has made drought adaptation and mitigation strategies a central focus in sustainable agricultural development.
Drought stress negatively affects a range of plant morphological, physiological, and biochemical processes, ultimately suppressing growth and development. Morphologically, drought-stressed plants often display stunted growth, reduced leaf area, and decreased stem diameter due to impaired cell expansion and limited water and nutrient translocation [5-8]. Physiologically, drought interferes with water uptake and transport, leading to reduced stomatal conductance, lowered transpiration rates, and diminished photosynthetic efficiency. Although many crop species have evolved tolerance mechanisms—such as stomatal closure and root elongation—these adaptations often come with trade-offs in biomass accumulation and reproductive output [9-10]. On the biochemical level, drought stress triggers the accumulation of osmoprotective compounds like proline, an increase in antioxidant enzyme activity, and elevated levels of secondary metabolites such as phenolics and flavonoids [11-14]. These biochemical adjustments help maintain cellular homeostasis and mitigate oxidative stress. However, the overall effectiveness of these morphological, physiological, and biochemical responses depends largely on the severity and duration of the stress, as well as the plant genotype. Amelioration strategies aimed at enhancing plant resilience to abiotic stress have received increasing attention in recent years. Among these, the use of organic soil amendments—also referred to as organic ameliorants—has been proposed as an eco-friendly and sustainable approach to improve soil fertility, water retention, and microbial activity [15]. Organic ameliorants such as compost, bokashi, animal manure, and biochar have been shown to enhance plant performance under stress conditions by improving soil structure, increasing nutrient availability, and stimulating beneficial soil microbiota. Bokashi may enhance microbial activity and nutrient availability, while biochar increases soil porosity and water retention, potentially leading to synergistic effects [16]. In particular, biochar has been noted for its high surface area and porosity, which contribute to its water-holding and cation exchange capacities, while bokashi—a fermented organic matter—can supply a wide range of nutrients and microbial inoculants [15, 16].
Despite growing evidence of the benefits of individual organic amendments, limited research has been conducted on their combined effects, especially under drought stress conditions. Furthermore, there is a lack of comparative studies that evaluate the synergistic or antagonistic interactions between different types of organic ameliorants. It remains unclear whether specific combinations of organic inputs may provide enhanced benefits for plant stress tolerance compared to their individual application. Understanding these interactions is crucial for developing integrated soil management practices that are both cost-effective and environmentally sustainable.
Watermelon (Citrullus lanatus L.) is a widely cultivated fruit crop known for its high water requirement and sensitivity to water scarcity. Drought stress during the vegetative or reproductive stages of watermelon significantly reduces fruit size, sugar content, and overall yield. As a result, effective drought mitigation strategies are essential for maintaining production in water-limited environments. Given the crop’s vulnerability, watermelon provides an ideal model for evaluating the role of organic ameliorants in alleviating abiotic stress.
This study aims to investigate the efficacy of different combinations of water hyacinth bokashi, manure, and rice husk biochar in mitigating the adverse effects of drought stress on watermelon. A factorial randomized block design was used to examine the interaction between four organic ameliorant combinations and four drought stress levels based on field capacity (FC). The research focused on assessing key biochemical and physiological parameters, including chlorophyll content, relative water content, proline accumulation, antioxidant activity, and total phenolic content, as indicators of plant response to drought. The findings are expected to contribute valuable insights into the development of sustainable soil amendment practices to enhance crop resilience under water-limited conditions.
2.1 Research location and time
This research was conducted in a controlled greenhouse environment from February to June 2024. The study site is located at an altitude of 5.5 meters above sea level (masl), positioned at coordinates 6°51′–7°23′ South Latitude and 112°33′–112°34′ East Longitude. The soil type at the location is predominantly clay. The plant material used was watermelon (Citrullus lanatus L.) variety Golden Inden-F1. Golden Inden-F1 was selected for its commercial relevance and sensitivity to water deficit, making it suitable for drought stress studies. Organic ameliorants used in this study included water hyacinth bokashi, livestock manure, and rice husk biochar. Water hyacinth bokashi was prepared through anaerobic fermentation for 28 days. Rice husk biochar was produced at 550℃ for 9 hours using a limited oxygen kiln.
2.2 Experimental design
The experiment was arranged using a Factorial Randomized Block Design (FRBD) with two factors and three replications to evaluate the combined effects of organic ameliorants and drought stress levels on watermelon growth. The first factor consisted of four combinations of organic ameliorants: (1) water hyacinth bokashi + manure (BoM), (2) water hyacinth bokashi + rice husk biochar (BoBi), (3) manure + rice husk biochar (MBi), and (4) a complete combination of water hyacinth bokashi, manure, and rice husk biochar (BoMBi). The second factor involved four levels of drought stress, defined by soil moisture availability at 100%, 75%, 50%, and 25% of field capacity (FC). Each treatment combination was replicated three times, with five sample plants per replication.
2.3 Preliminary study
A preliminary study was conducted to determine the field capacity (FC) threshold for watermelon growth under drought stress. Watermelon was planted in polybags (40 cm × 50 cm) containing each ameliorant combination, with six replications per treatment. Plants were irrigated daily for three weeks to establish growth, followed by water withdrawal until visual symptoms of temporary wilting appeared. Temperatures ranged from 29–35℃ with 65–80% relative humidity. Drought was simulated by controlling irrigation based on gravimetric soil moisture to reach 100%, 75%, 50%, and 25% FC. Soil samples were then collected and analyzed to determine the moisture content at the wilting point. Based on these observations, 25% FC was identified as the critical threshold for drought stress response in watermelon.
2.4 Preparation of planting media
Planting media consisted of a 1:1 ratio mixture of clay soil and organic ameliorant according to each treatment. The clay soil used had a pH of 6.3 and 2.1% organic matter content, which may influence water retention and amendment efficacy. Media were filled into 40 cm × 50 cm polybags and arranged in the greenhouse with 60 cm × 70 cm spacing. All polybags were initially irrigated to 100% FC and allowed to stabilize for three weeks. Furadan 3G insecticide was applied to the media one day before transplanting to prevent soil-borne pests.
2.5 Planting, watering, and harvesting procedure
Watermelon seeds were germinated and maintained until they reached the transplanting stage. One healthy seedling was transplanted into each polybag. Each treatment combination included three replicates, with five plants per replicate.
Irrigation was carried out according to the designated drought stress treatments using the gravimetric method. Soil samples were taken at a depth of 20 cm to determine moisture content by oven-drying. Water was added based on the difference between current moisture levels and the target FC, with irrigation conducted either in the morning or evening as required.
Harvesting was performed once fruits reached physiological maturity, identified by several visual and tactile indicators: clean and shiny skin, browning of tendrils at the fruit base, dull sound when tapped, and yellowing of the underside of the fruit. In addition, the stalks showed signs of aging and detachment readiness.
2.6 Observational parameters and data analysis
Observations were conducted on both agronomic and physiological characteristics of watermelon plants. Agronomic parameters included root length and root dry weight, measured destructively at harvest. Roots were oven-dried at a temperature of 70–80℃ for 48 hours to determine their dry weight. Physiological parameters included chlorophyll a, b, and total chlorophyll contents, measured at 4, 6, and 8 weeks after planting using spectrophotometry. Leaf relative water content (RWC) was calculated using the method of Barrs and Weatherley, where RWC (%) = [(FW − DW)/(SW − DW)] × 100, with FW as fresh weight, DW as dry weight, and SW as saturated weight. Proline content was estimated through acid ninhydrin extraction, while antioxidant activity was determined using the DPPH (2,2-diphenyl-1-picrylhydrazyl) assay. Total phenolic content (TPC) was analyzed using the Folin-Ciocalteu method. In addition, fruit sweetness was measured using a hand-held refractometer and expressed in degrees Brix (°Brix). All data were subjected to analysis of variance (ANOVA) using a one-way factorial design. Treatment means were compared using Duncan’s Multiple Range Test (DMRT) at a significant level of p < 0.05 to determine the effect of drought stress and ameliorant combinations on measured variables.
This section presents a comprehensive evaluation of the physiological responses of watermelon plants subjected to varying levels of drought stress and the mitigating effects of different combinations of organic ameliorants. The parameters analyzed include root length, proline content, antioxidant activity, chlorophyll content, and relative water content (RWC) of leaves.
3.1 Root length
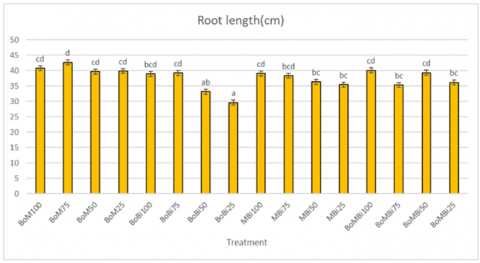
Root length was measured destructively at harvest. The shortest root length was observed under the BoBi25 treatment (water hyacinth bokashi + rice husk biochar under 25% FC drought stress), while the BoM treatment (bokashi + manure) showed relatively stable root length across drought levels (see Figure 1).
Roots play a pivotal role in plant survival under drought by enabling water and nutrient uptake and triggering adaptive responses [17]. Plants exposed to drought often respond by developing deeper or more extensive root systems to access moisture from deeper soil layers [18]. However, the presence of high water-retentive ameliorants like biochar and bokashi may reduce the plant's need to elongate roots, resulting in shorter root systems in high-stress conditions. The ability of BoBi to retain water close to the root zone likely minimized the necessity for deeper root growth.
3.2 Proline content
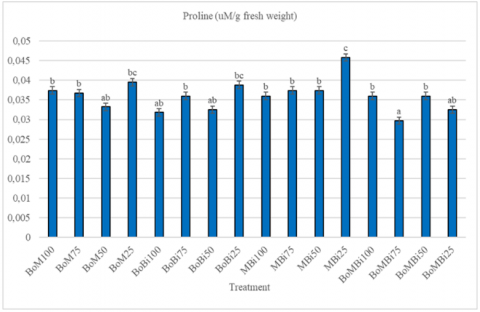
Proline is an amino acid that plants produce when experiencing abiotic stress such as drought. Several studies have stated that proline is reported to confer plant resistance to drought stress. Proline accumulation is a common response to abiotic stress and serves as an osmoprotectant. The highest proline levels, as can be seen in Figure 2, were recorded in the MBi25 treatment (manure + biochar under 25% FC stress), indicating a significant stress response.
In a drought-stress-prone environment, plants may adapt by accumulating proline. These results show that the combination of manure and biochar is less able to maintain soil moisture than other treatments. Therefore, watermelon growth in media can be maintained with the addition of an ameliorant by producing high proline [19]. Water potential decreases with increased proline content, and osmolytes accumulate during the osmoregulation process, allowing plants to absorb water for growth and metabolic processes [20-22].
Increased proline accumulation is indicative of plant adaptation under stress, as it supports osmotic balance, stabilizes proteins and membranes, and scavenges reactive oxygen species [19, 20]. The elevated proline content in MBi25 suggests that this combination was less effective at conserving water compared to others, triggering a stronger biochemical response. These findings are consistent with studies reporting a positive correlation between drought tolerance and proline biosynthesis. Plants survive drought stress by using biochemical, structural, and molecular defense mechanisms, including the formation of osmolytes such as proline, proteins, carbohydrates, and glycine betaine. Additionally, the use of salicylic acid increases resistance to drought stress by maintaining the redox potential and triggering proline biosynthesis [23, 24].
3.3 Antioxidant activity
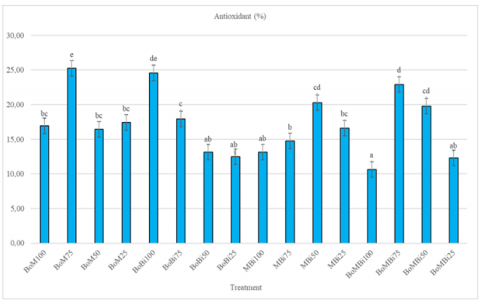
Drought stress is known to trigger the excessive production of Reactive Oxygen Species (ROS), which can lead to cellular damage if not properly managed. To counteract this, plants activate antioxidant defense systems comprising both enzymatic and non-enzymatic components [24, 25]. These mechanisms are crucial for maintaining cellular homeostasis, mitigating oxidative stress, and supporting survival under adverse environmental conditions. In this study, antioxidant activity varied depending on the drought intensity and ameliorant treatment (Figure 3). The highest antioxidant levels were observed under BoM75 (25.25%), BoBi100 (24.58%), and BoMBi75 (22.92%) treatments, indicating that mild to moderate drought stress in combination with organic amendments can stimulate antioxidant defenses.
Conversely, antioxidant levels declined under severe drought stress (25% FC), particularly in BoBi25 and BoMBi25 treatments, which recorded the lowest values. This suggests a physiological trade-off under extreme stress, where the plant may divert metabolic energy from antioxidant synthesis toward vital survival functions such as osmolyte accumulation or stress signaling. Despite this reduction, the plant's resilience indicates an efficient adaptive strategy, allowing it to maintain essential physiological processes without requiring substantially elevated antioxidant levels [26, 27]. Furthermore, gene expression of antioxidant-related enzymes is often modulated by stress severity, species-specific traits, developmental stage, and metabolic status, which could explain the treatment-dependent responses observed in this study.
Several plant species have demonstrated increased enzymatic antioxidant activity in response to water deficit conditions. Notably, enhanced antioxidant responses have been observed in barley (Hordeum vulgare L.) [24], beans [28], maize [29], wheat [27, 30], and rice [31]. These findings underscore the vital role of both enzymatic and non-enzymatic antioxidant defense systems in mitigating the adverse effects of drought stress. By detoxifying and scavenging excess Reactive Oxygen Species (ROS), these mechanisms protect cellular structures, maintain metabolic stability, and ultimately enhance plant resilience under drought conditions.
3.4 Chlorophyll content
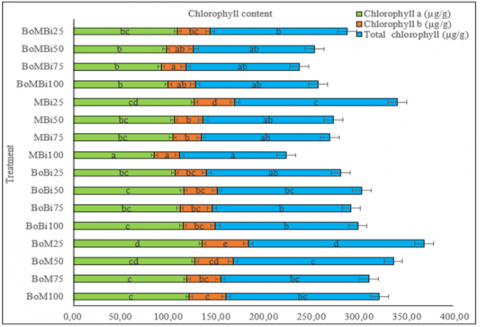
Chlorophyll degradation is a typical physiological response to drought stress, often resulting in diminished photosynthetic efficiency and reduced biomass accumulation. As drought severity increases, chlorophyll content and leaf area usually decline, thereby affecting photosynthetic activity, organic matter accumulation, and overall plant growth [32]. However, in this study, a contrasting trend was observed—plants subjected to moderate and severe drought conditions (25% and 50% field capacity), particularly under MBi25, BoM25, and BoM50 treatments, maintained or even enhanced their chlorophyll content compared to some well-watered controls (Figure 4). Specifically, chlorophyll a content reached 127.57 µg/g (MBi25), 135.84 µg/g (BoM25), and 128.09 µg/g (BoM50); chlorophyll b content was 42.40 µg/g, 48.29 µg/g, and 39.91 µg/g, respectively. Total chlorophyll content in these treatments ranged from 168.00 to 184.13 µg/g.
This unexpected increase suggests a potential protective or compensatory mechanism facilitated by the organic ameliorants, allowing the plants to maintain chlorophyll biosynthesis and photosynthetic efficiency even under water-limited conditions. These findings align with previous studies indicating that certain stress-adaptive responses, such as osmolyte accumulation and enhanced antioxidant activity, may help stabilize chlorophyll levels during drought [33]. The results imply that strategic combinations of organic ameliorants can play a critical role in mitigating drought-induced chlorophyll loss and supporting physiological resilience in watermelon.
3.5 Leaf relative water content (RWC)
RWC is a widely recognized and reliable indicator of the water status within plant tissues, reflecting a plant’s ability to maintain hydration and tolerate dehydration under stress conditions. It provides an integrative measure of both leaf water deficit and the severity of environmental stress caused by factors such as drought and heat. RWC is influenced by water potential and cellular mechanisms that regulate water retention and maintain structural integrity. In addition to its role in assessing water balance, RWC also reflects the plant’s capacity to absorb and mitigate the effects of harmful substances in the soil, such as salts or heavy metals [34]. Biochar exerted a direct physical effect by improving water retention, while bokashi contributed indirectly via microbial activity that enhanced nutrient cycling and stress signaling pathways. Plants that can sustain higher RWC under stress maintain cell turgor more effectively, giving them a physiological advantage for continued growth and function in adverse conditions. As such, RWC serves as a key physiological parameter in evaluating a plant's drought tolerance and overall resilience in extreme environments.
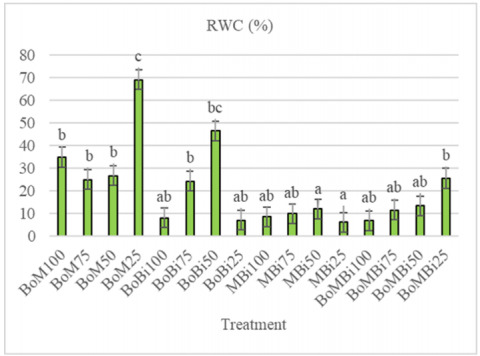
The highest RWC was obtained by BoM25 and BoBi50 treatments at 69.70% and 46.46%, as shown in Figure 5. In this context, bokashi ameliorant combined with biochar and manure can store water. The high RWC values in these treatments suggest that combinations of bokashi with manure or biochar improved water retention and mitigated drought stress. These amendments enhance soil porosity, water-holding capacity, and nutrient availability, thereby supporting better water uptake and retention by the plant [34]. Coupled with the accumulation of osmolytes such as proline, these mechanisms contribute to osmotic adjustment and sustained cell turgor under drought conditions. The co-occurrence of high chlorophyll and high proline in MBi25 suggests a ‘high-cost tolerance’ strategy, where physiological functioning is preserved under stress through elevated biochemical adaptation.
Watermelon is considered a drought-resilient crop due to its ability to activate specific physiological and biochemical adaptations that help maintain or enhance RWC under water-limited conditions. These adaptive mechanisms include improved water use efficiency and modifications in root architecture that facilitate more effective water uptake from deeper or drier soil layers. In this study, treatments such as BoM25 and MBi25 exhibited notably high RWC values, which correlated with elevated proline accumulation. The accumulation of osmolytes—including proline, glycine betaine, and soluble sugars—plays a crucial role in maintaining cellular osmotic balance. These compounds help preserve cell turgor by stabilizing osmotic pressure and retaining water within cells, even under conditions of external water scarcity. The positive correlation between chlorophyll retention and RWC aligns with findings in maize [35]. Moreover, elevated proline levels under MBi25 support previous reports of osmolyte-driven drought adaptation in legumes [36].
3.6 Correlation between chlorophyll decline, water relations, and sweetness accumulation
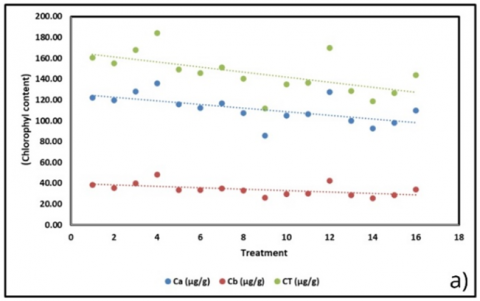
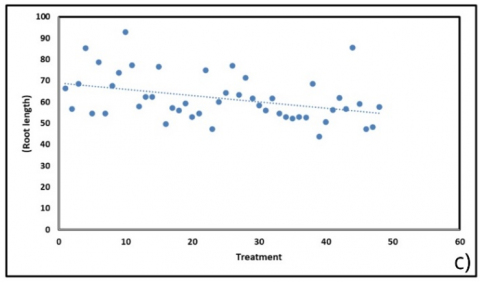
The correlation analysis revealed that substrate treatments exerted significant yet varying influences on plant physiological and biochemical parameters (Figure 6). Chlorophyll content (Figure 6(a)), particularly chlorophyll a (Ca), showed the strongest negative trend, decreasing by approximately 1.74 µg/g per treatment change (R² = 0.372). This pattern was consistent with chlorophyll b (–0.68 µg/g; R² = 0.287) and total chlorophyll (–2.42 µg/g; R² = 0.351), suggesting that more complex substrate mixtures diminished pigment accumulation. The reduction in chlorophyll content was also associated with a decline in root length (Figure 6(c); –0.30 units; R² = 0.146), which may reflect impaired photosynthetic efficiency leading to reduced carbon allocation to root development.
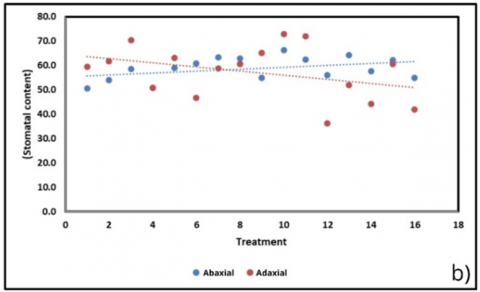
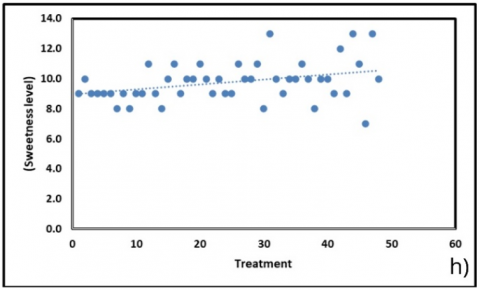
Stomatal traits (Figure 6(b)) displayed divergent responses, with abaxial density increasing slightly (+0.40 units; R² = 0.157) while adaxial density declined (–0.84 units; R² = 0.133). The balance between these two traits may represent a compensatory mechanism: as adaxial stomata decreased, abaxial stomata increased, thereby maintaining gas exchange. Interestingly, this adjustment correlated with a modest but consistent increase in sweetness (Figure 6(h); +0.033 °Brix per treatment; R² = 0.124). The upward trend in sweetness, despite declining chlorophyll, suggests that carbohydrate partitioning remained stable, possibly supported by enhanced abaxial stomatal activity that sustained photosynthetic carbon assimilation under suboptimal pigment conditions.
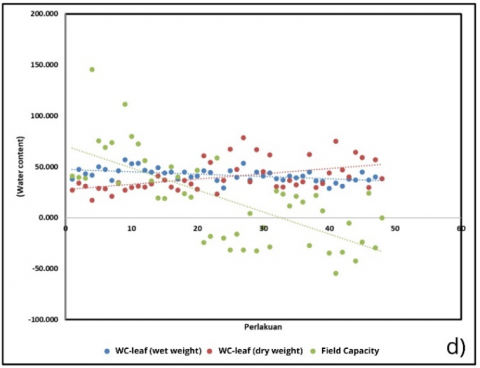
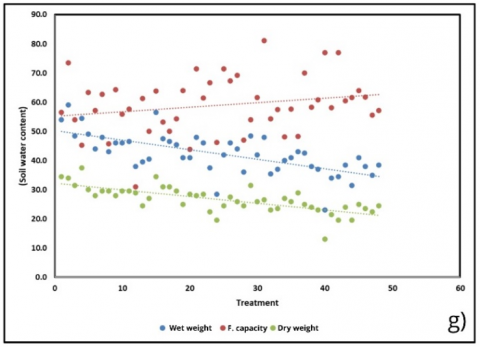
Water relations further clarified treatment effects. Fresh weight water content declined (–0.23 units; R² = 0.272), whereas dry weight water content increased (+0.52 units; R² = 0.235), indicating a shift in tissue hydration and solute concentration (Figure 6(d)). These physiological responses aligned with soil water parameters (Figure 6(g)), where fresh soil moisture decreased (–0.33 units; R² = 0.443) and dry weight soil content showed the strongest association (–0.23 units; R² = 0.517). The pronounced decline in field capacity (–2.16 units; R² = 0.498) paralleled reductions in chlorophyll, reinforcing the link between substrate water-holding capacity and photosynthetic performance. The modest rise in sweetness (Figure 6(h)) may also be explained in this context, as lower water availability could promote solute concentration, resulting in higher °Brix readings.
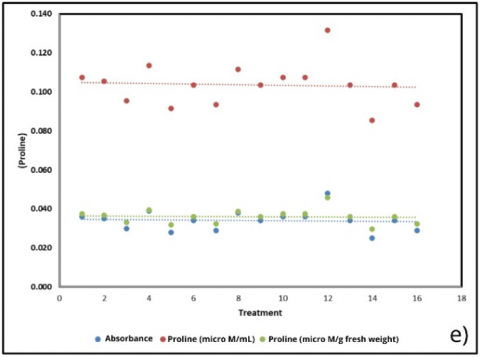
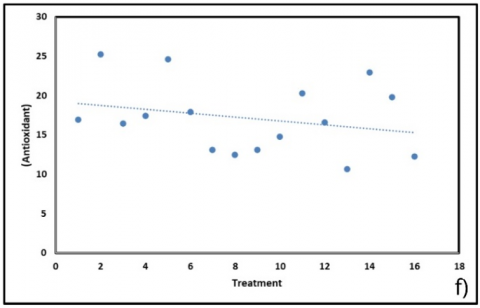
Figure 6. Correlation of physiological and biochemical parameters under substrate treatments (BoM = bokashi + manure; BoBi = bokashi + biochar; MBi = manure + biochar; BoMBi = bokashi + manure + biochar) (a) Chlorophyll content; (b) Stomatal content; (c) Root length; (d, g) Leaf and soil water relations; (e) Proline; (f) antioxidant activity; (h) Sweetness (°Brix) level
Proline accumulation (Figure 6(e)), commonly a stress marker, showed negligible correlation with treatments (R² ≈ 0.004), indicating that plants did not experience substantial osmotic stress despite measurable changes in water balance. Similarly, antioxidant activity (Figure 6(f)) exhibited only a weak negative association (–0.25 units; R² = 0.071), suggesting that oxidative stress was not a dominant factor under the tested conditions. Together, the absence of strong proline and antioxidant responses suggests that treatment effects were mediated primarily through changes in structural water relations and resource allocation rather than stress-induced metabolic pathways.
In summary, the strongest treatment effects were observed in chlorophyll dynamics (Figure 6(a)) and soil water parameters (Figure 6(g)), both of which play central roles in determining plant productivity. Stomatal compensation (Figure 6(b)) and the modest but consistent increase in sweetness (Figure 6(h)) indicate adaptive adjustments that allowed carbohydrate accumulation to persist, even under reduced pigment levels and limited water availability. By contrast, stress-related metabolites such as proline (Figure 6(e)) and antioxidants (Figure 6(f)) remained largely unaffected, suggesting that the physiological shifts were structural and metabolic rather than stress-driven. This contrast provides important context for the present findings. In the current study, watermelon grown under bokashi–manure–biochar amendments exhibited significant declines in chlorophyll and water parameters (Figure 6(a, c, d, g)), but these declines were not accompanied by strong proline accumulation (Figure 6(e)) or antioxidant activation (Figure 6(f)). Instead, adaptive responses were reflected in stomatal compensation and the modest increase in sweetness (Figure 6(b, h)), indicating that carbohydrate assimilation and solute concentration were maintained despite pigment and water loss. This aligns with earlier evidence that watermelon’s resilience relies more on structural and photosynthetic adjustments than on classical stress metabolites such as proline.
Comparative studies have shown that crops employ diverse physiological and biochemical strategies to cope with drought stress, and these strategies often differ markedly among species. In tobacco, the dominant response involves biochemical adjustments such as proline accumulation and the activation of antioxidant pathways [37]. Proline serves as a critical osmolyte that facilitates osmotic adjustment, stabilizes proteins and membranes, and mitigates oxidative stress by scavenging reactive oxygen species (ROS) [38, 39]. Similarly, enhanced antioxidant activity of crops functions as a front-line defense against ROS, enabling these species to withstand prolonged periods of water deficit [40, 41]. However, these adjustments are often accompanied by degradation of chlorophyll pigments, where drought-induced gene expression triggers the breakdown of photosynthetic pigments despite the application of soil amendments [42, 43].
By contrast, watermelon (Citrullus lanatus) demonstrates a distinctive response when subjected to water stress in the presence of organic amendments such as bokashi–manure combinations. Rather than relying primarily on osmolyte accumulation or antioxidant pathways, watermelon exhibits a capacity to stabilize chlorophyll content under stress [44, 45]. This stabilization is likely mediated by amendment-induced activation of stress-responsive genes related to photosynthesis and antioxidative defense, which collectively preserve pigment integrity and sustain photosynthetic activity [46, 47]. Such responses highlight the central role of chlorophyll maintenance in watermelon’s drought resilience, in contrast to the more biochemical-centric strategies observed in peanuts and grasses.
Taken together, these comparisons underscore crop-specific strategies in drought adaptation. Watermelon’s strategy emphasizes the stabilization of chlorophyll pigments and maintenance of carbohydrate metabolism under organic amendment regimes. These differences emphasize the need for crop-tailored agronomic interventions: organic amendments may be especially effective for watermelon, while biochemical fortification strategies may be more relevant for other plants [48].
This study demonstrated that drought stress significantly impacts plant physiological responses and productivity, with watermelon plants exhibiting various adaptive mechanisms depending on stress severity. Among all treatments, BoM25 (bokashi + manure at 25% FC) was the most effective in maintaining high chlorophyll content (184.13 µg/g), RWC (69.70%), and physiological stability under severe drought. Therefore, BoM at 25% FC is recommended as the optimal combination for enhancing watermelon drought tolerance through organic amendment.
The author would like to thank the Indonesian Education Scholarship for the financial support provided to cover the costs of the education (SK No.: 03133/J5.2.3./BPI.06/10/2022/ 202231103879).
[1] Siriwach, R., Matsuda, F., Yano, K., Hirai, M.Y. (2020). Drought stress responses in context-specific genome-scale metabolic models of Arabidopsis thaliana. Metabolites, 10(4): 159. https://doi.org/10.3390/metabo10040159
[2] Madadgar, S., AghaKouchak, A., Farahmand, A., Davis, S.J. (2017). Probabilistic estimates of drought impacts on agricultural production. Geophysical Research Letters, 44(15): 7799-7807. https://doi.org/10.1002/2017gl073606
[3] Ullah, N., Ditta, A., Khalid, A., Mehmood, S., Rizwan, M.S., Ashraf, M., Iqbal, M.M. (2019). Integrated effect of algal biochar and plant growth promoting rhizobacteria on physiology and growth of maize under deficit irrigations. Journal of Soil Science and Plant Nutrition, 20(2): 346-356. https://doi.org/10.1007/s42729-019-00112-0
[4] Van Loon, A.F., Gleeson, T., Clark, J., Van Dijk, A.I.J.M., et al. (2016). Drought in the anthropocene. Nature Geoscience, 9(2): 89-91. https://doi.org/10.1038/ngeo2646
[5] Aslam M.M., Waseem, M., Jakada, B.H., Okal, E.J., Lei, Z., Saqib, H.S.A., Yuan, W., Xu, W., Zhang, Q. (2022). Mechanisms of abscisic acid-mediated drought stress responses in plant. International Journal of Molecular Sciences, 23(3): 1084. https://doi.org/10.3390/ijms23031084
[6] Yahaya, M.A., Shimelis, H. (2022). Drought stress in sorghum: Mitigation strategies, breeding methods and technologies—A review. Journal of Agronomy and Crop Science, 208(2): 127-142. https://doi.org/10.1111/jac.12573
[7] Queiroz, M.S., Oliveira, C.E.S., Steiner, F., Zuffo, A.M., Zoz, T., Vendruscolo, E.P., Silva, M.V., Mello, B.F.F.R., Cabral, R.C., Menis, F.T. (2019). Drought stresses on seed germination and early growth of maize and sorghum. Journal of Agricultural Science, 11(2): 310. https://doi.org/10.5539/jas.v11n2p310
[8] Wagaw, K. (2019). Review on mechanisms of drought tolerance in sorghum (Sorghum bicolor (L.) Moench) basis and breeding methods. Academic Research Journal of Agricultural Science and Research, 7(2): 87-99. https://doi.org/10.14662/ARJASR2019.007
[9] Pepe, M., Crescente, M.F., Varone, L. (2022). Effect of water stress on physiological and morphological leaf traits: A comparison among the three widely-spread invasive alien species Ailanthus altissima, Phytolacca americana, and Robinia pseudoacacia. Plant, 11(7): 899. https://doi.org/10.3390/plant11070899
[10] McDowell, N.G., Ball, M., Bond-Lamberty, B., Kirwan, M.L., Krauss, K.W., Megonigal, J.P., Mencuccini, M., Ward, N.D., Weintraub, M.N., Bailey, V. (2022). Processes and mechanisms of coastal woody-plant mortality. Global Change Biology, 28(20): 5881-5900. https://doi.org/10.1111/gcb.16297
[11] Gao, J., Zhang, R.H., Wang, W.B., Li, Z.W., Xue, J.Q. (2015). Effects of drought stress on performance of photosystem II in maize seedling stage. The Journal of Applied Ecology, 26(5): 1391-1396.
[12] Islam, M.R., Sarker, B.C., Alam, M.A., Javed, T., Alam, M.J., Zaman, M.S.U., Azam, M.G., Shabbir, R., Raza, A., Habib-ur-Rahman, M., Dessoky, E.S., Islam, M.S. (2021). Yield stability and genotype environment interaction of water deficit stress tolerant mung bean (Vigna radiata L. Wilczak) genotypes of Bangladesh. Agronomy, 11(11): 2136. https://doi.org/10.3390/agronomy11112136.
[13] Mohagheghian, B., Saeidi, G., Arzani, A. (2025). Phenolic compounds, antioxidant enzymes, and oxidative stress in barley (Hordeum vulgare L.) genotypes under field drought-stress conditions. BMC Plant Biology, 25: 709. https://doi.org/10.1186/s12870-025-06750-0
[14] Akhtar, G., Faried, H.N., Razzaq, K., Ullah, S., et al. (2022). Chitosan-induced physiological and biochemical regulations confer drought tolerance in pot marigold (Calendula officinalis L.). Agronomy, 12(2): 474. https://doi.org/10.3390/agronomy12020474
[15] Rashad, R.T., El-Agyzy, F.H.A., Abdel-Azeem, S.M. (2018). Impact of irrigation intervals on the yield and quality of lupine (Lupinus termis L.) grown in sandy soil amended by an organic amendment. Asian Soil Research Journal, 1(3): 1-11. https://doi.org/10.9734/asrj/2018/v1i3693.
[16] Abd El-Mageed, T.A., El-Sherif, A.M.A., Abd El-Mageed, S.A., Abdou, N.M. (2019). A novel compost alleviate drought stress for sugar beet production grown in Cd-contaminated saline soil. Agricultural Water Management, 226: 105831. https://doi.org/10.1016/j.agwat.2019.105831
[17] Shoaib, M., Banerjee, B.P., Hayden, M., Kant, S. (2022). Roots' drought adaptive traits in crop improvement. Plants, 11(17): 2256. https://doi.org/10.3390/plants11172256
[18] Ranjan, A., Sinha, R., Singla-Pareek, S.L., Pareek, A., Singh, A.K. (2022). Shaping the root system architecture in plants for adaptation to drought stress. Physiologia Plantarum, 174(2): e13651. https://doi.org/10.1111/ppl.13651
[19] Doneva, D., Pál, M., Brankova, L., Szalai, G., Tajti, J., Khalil, R., Ivanovska, B., Velikova, V., Misheva, S., Janda, T., Peeva, V. (2021). The effects of putrescine pre-treatment on osmotic stress responses in drought-tolerant and drought-sensitive wheat seedlings. Physiologia Plantarum, 171(2): 200-216. https://doi.org/10.1111/ppl.13150
[20] Shahid, M.A., Sarkhosh, A., Khan, N., Balal, R.M., Ali, S., Rossi, L., Gómez, C., Mattson, N., Nasim, W., Garcia-Sanchez, F. (2020). Insights into the physiological and biochemical impacts of salt stress on plant growth and development. Agronomy, 10(7): 938. https://doi.org/10.3390/agronomy10070938
[21] Iqbal, M.J. (2018). Role of osmolytes and antioxidant enzymes for drought tolerance in wheat. In Global Wheat Production, 51: 51-66. https://doi.org/10.5772/intechopen.75926
[22] El-Beltagi, H.S., Mohamed, H.I., Sofy, M.R. (2020). Role of ascorbic acid, glutathione and proline applied as singly or in sequence combination in improving chickpea plant through physiological change and antioxidant defense under different levels of irrigation intervals. Molecules, 25(7): 1702. https://doi.org/10.3390/molecules25071702
[23] Sytar, O., Kumari, P., Yadav, S., Brestic, M., Rastogi, A. (2019). Phytohormone priming: Regulator for heavy metal stress in plant. Journal of Plant Growth Regulation, 38(2): 739-752. https://doi.org/10.1007/s00344-018-9886-8
[24] Abdelaal, K.A.A., Attia, K.A., Alamery, S.F., El-Afry, M.M., Ghazy, A.I., Tantawy, D.S., Al-Doss, A.A., El-Shawy, E.S.E., Abu-Elsaoud, A.M., Hafez, Y.M. (2020). Exogenous application of proline and salicylic acid can mitigate the injurious impacts of drought stress on barley plant associated with physiological and histological characters. Sustainability, 12(5): 1736. https://doi.org/10.3390/su12051736
[25] Kumar, R., Pareek, N.K., Kumar, U., Javed, T., et al. (2022). Coupling effects of nitrogen and irrigation levels on growth attributes, nitrogen use efficiency, and economics of cotton. Frontiers in Plant Science, 13: 890181. https://doi.org/10.3389/fpls.2022.890181
[26] Chiappero, J., Cappellari, L.D.R., Alderete, L.G.S., Palermo, T.B., Banchio, E. (2019). Plant growth promoting rhizobacteria improve the antioxidant status in Mentha piperita grown under drought stress leading to an enhancement of plant growth and total phenolic content. Industrial Crops and Products, 139: 111553. https://doi.org/10.1016/j.indcrop.2019.111553
[27] Vuković, R., Čamagajevac, I.Š., Vuković, A., Šunić, K., Begović, L., Mlinarić, S., Sekulić, R., Sabo, N., Španić, V. (2022). Physiological, biochemical and molecular response of different winter wheat varieties under drought stress at germination and seedling growth stage. Antioxidants, 11(4): 693. https://doi.org/10.3390/antiox11040693
[28] Nawaz, M., Wang, X., Saleem, M.H., Khan, M.H.U., Afzal, J., Fiaz, S., Ali, S., Ishaq, H., Khan, A.H., Rehman, N., Shaukat, S., Ali, S. (2021). Deciphering Plantago ovata forsk leaf extract mediated distinct germination, growth and physio-biochemical improvements under water stress in maize (Zea mays L.) at early growth stage. Agronomy, 11(7): 1404. https://doi.org/10.3390/agronomy11071404
[29] Shafiq, S., Akram, N.A., Ashraf, M., García-Caparrós, P., Ali, O.M., Latef, A.A.H.A. (2021). Influence of glycine betaine (natural and synthetic) on growth, metabolism and yield production of drought-stressed maize (Zea mays L.) Plant. Plant, 10(11): 2540. https://doi.org/10.3390/plant10112540
[30] Mustafa, H., Ilyas, N., Akhtar, N., Raja, N.I., Zainab, T., Shah, T., Ahmad, A., Ahmad, P. (2021). Biosynthesis and characterization of titanium dioxide nanoparticles and its effects along with calcium phosphate on physicochemical attributes of wheat under drought stress. Ecotoxicology and Environmental Safety, 223: 112519. https://doi.org/10.1016/j.ecoenv.2021.112519
[31] Wang, X., Liu, H., Yu, F., Hu, B., Jia, Y., Sha, H., Zhao, H. (2019). Differential activity of the antioxidant defence system and alterations in the accumulation of osmolyte and reactive oxygen species under drought stress and recovery in rice (Oryza sativa L.) tillering. Scientific Reports, 9(1): 8543. https://doi.org/10.1038/s41598-019-44958-x
[32] He, H., Wang, Q., Wang, L., Yang, K., Yang, R., You, C., Ke, J., Wu, L. (2021). Photosynthetic physiological response of water-saving and drought-resistant rice to severe drought under wetting-drying alternation irrigation. Physiologia Plantarum, 173(4): 2191-2206. https://doi.org/10.1111/ppl.13568
[33] Sherin, G., Aswathi, K.P.R., Puthur, J.T. (2022). Photosynthetic functions in plant subjected to stresses are positively influenced by priming. Plant Stress, 4: 100079. https://doi.org/10.1016/j.stress.2022.100079
[34] Raklami, A, Meddich, A, Oufdou, K, Baslam, M. (2022). Plants-Microorganisms-based bioremediation for heavy metal cleanup: Recent developments, phytoremediation techniques, regulation mechanisms, and molecular responses. International Journal of Molecular Sciences, 23(9): 5031. https://doi.org/10.3390/ijms23095031
[35] Badr, A., Brüggemann, W. (2020). Comparative analysis of drought stress response of maize genotypes using chlorophyll fluorescence measurements and leaf relative water content. Photosynthetica, 58(SI): 638-645. https://doi.org/10.32615/ps.2020.014
[36] Noor, M.M.A., Tahjib-Ul-Arif, M., Alim, S.M.A., Islam, M.M., Hasan, M.T., Babar, M.A., Hossain, M.A., Jewel, Z.A., Murata, Y., Mostofa, M.G. (2024). Lentil adaptation to drought stress: Response, tolerance, and breeding approaches. Frontiers in Plant Science, 15: 1403922. https://doi.org/10.3389/fpls.2024.1403922
[37] Khan, R., Ma, X., Zhang, J., Wu, X., Iqbal, A., Wu, Y., Wang, S. (2021). Circular drought-hardening confers drought tolerance via modulation of the antioxidant defense system, osmoregulation, and gene expression in tobacco. Physiologia Plantarum, 172(2): 1073-1088. https://doi.org/10.1111/ppl.13402
[38] Zhang, Y., Yang, S., Dao, J., Deng, J., Shahzad, A., Fan, X., Zeng, Z. (2020). Drought-induced alterations in photosynthetic, ultrastructural and biochemical traits of contrasting sugarcane genotypes. Plos One, 15(7): e0235845. https://doi.org/10.1371/journal.pone.0235845
[39] Hasanuzzaman, M., Bhuyan, M.H.M.B., Zulfiqar, F., Raza, A., Mohsin, S., Mahmud, J., Fotopoulos, V. (2020). Reactive oxygen species and antioxidant defense in plants under abiotic stress: Revisiting the crucial role of a universal defense regulator. Antioxidants, 9(8): 681. https://doi.org/10.3390/antiox9080681
[40] Ahmad, S., Kamran, M., Ding, R., Meng, X., Wang, H., Ahmad, I., Fahad, S., Han, Q. (2019). Exogenous melatonin confers drought stress by promoting plant growth, photosynthetic capacity and antioxidant defense system of maize seedlings. PeerJ, 7: e7793. https://doi.org/10.7717/peerj.7793
[41] Ye, L., Zhao, X., Bao, E., Cao, K., Zou, Z. (2019). Effects of arbuscular mycorrhizal fungi on watermelon growth, elemental uptake, antioxidant, and photosystem II activities and stress-response gene expressions under salinity-alkalinity stresses. Frontiers in Plant Science, 10. https://doi.org/10.3389/fpls.2019.00863
[42] Azeem, A., Mai, W., Gul, B., Rasheed, A. (2025). Influence of soil amendment application on growth and yield of Hedysarum scoparium Fisch. et mey and Avena sativa L. under saline conditions in dry-land regions. Plants, 14(6): 855. https://doi.org/10.3390/plants14060855
[43] Hoque, M.N., Imran, S., Hannan, A., Paul, N.C., Mahamud, M.A., Chakrobortty, J., Sarker, P., Irin, I.J., Brestic, M., Rhaman, M.S. (2022). Organic amendments for mitigation of salinity stress in plants: A review. Life, 12(10): 1632. https://doi.org/10.3390/life12101632
[44] Erez, M.E., İnal, B., Karipçin, M.Z., Altıntaş, S. (2020). Physiological and gene-expression variations in watermelon (Citrullus lanatus L.) cultivars exposed to drought stress. Acta Societatis Botanicorum Poloniae, 89(2). https://doi.org/10.5586/asbp.8921
[45] Abdelaziz, M.E., Atia, M.A.M., Abdelsattar, M., Abdelaziz, S.M., Ibrahim, T.A.A., Abdeldaym, E.A. (2021). Unravelling the role of Piriformospora indica in combating water deficiency by modulating physiological performance and chlorophyll metabolism-related genes in Cucumis sativus. Horticulturae, 7(10): 399. https://doi.org/10.3390/horticulturae7100399
[46] Malambane, G., Madumane, K., Sewelo, L., Batlang, U. (2023). Drought stress tolerance mechanisms and their potential common indicators to salinity, insights from the wild watermelon (Citrullus lanatus): A review. Frontiers in Plant Science, 13: 1074395. https://doi.org/10.3389/fpls.2022.1074395
[47] Mendoza-Labrador, J., Perdomo, F., Hernández, J., Uribe, D., Buitrago, R. (2019). Enhancement of drought tolerance on guinea grass by dry alginate macrobeads as inoculant of Bacillus strains. https://doi.org/10.1101/761056
[48] El-Katony, T.M., Ward, F.M., Deyab, M.A., El-Adl, M.F. (2021). Algal amendment improved yield and grain quality of rice with alleviation of the impacts of salt stress and water stress. Heliyon, 7(9): e07911. https://doi.org/10.1016/j.heliyon.2021.e07911