FimH36-kDa Salmonella Typhi Protein as an Inhibitor of Enteric Pathogen Adhesion for the Development of Biological Material for a Typhoid Vaccine

I Nengah Kundera*![]() | Maharani Ajeng Astiti
| Maharani Ajeng Astiti![]() | Raya Agni
| Raya Agni![]() | Achmad Ramadhan
| Achmad Ramadhan![]()
© 2025 The authors. This article is published by IIETA and is licensed under the CC BY 4.0 license (http://creativecommons.org/licenses/by/4.0/).
OPEN ACCESS
The adherence of Salmonella enterica serotype typhi to the surface of intestinal cells is an important step in infection, which is facilitated by fimbriae. This study aims to evaluate the inhibitory effects of the FimH36-kDa protein from the S. Typhi fimbriae on the adhesion of Escherichia coli, Shigella flexneri, and Salmonella Typhi to mouse enterocyte cells in vitro as part of the exploration of potential biological materials for typhoid vaccine development. This research method is exploratory and experimental through the isolation of the protein FimH36-kDa S. Typhi and the determination of its molecular weight. The FimH36-kDa protein was purified and characterized by SDS-PAGE and Western blot using AdhO36 monoclonal antibodies. Enterocyte models, derived from Mus musculus Balb/c intestinal cells and the Salmonella Typhi strain [8873], were used for in vitro adhesion assays. Inhibitory activity was assessed at 0–400 μg protein concentrations, with each dose applied to 300 μL of enterocyte suspension. Gram staining is performed on enterocytes, observed microscopically, and the adhesion index is calculated. The FimH36-kDa protein significantly reduces bacterial adhesion to enterocytes, with the highest inhibition observed in S. Typhi (92.86%), followed by S. flexneri (90.91%) and E. coli (85.4%). These results highlight its potential as a biological component candidate for an S. Typhi vaccine and as a protective agent against enteric pathogens.
bacterial adhesion, enterocyte, FimH36-kDa protein, Salmonella Typhi, vaccine
Adhesion and invasion by pathogenic bacteria represent critical early events in the pathogenesis process. In S. Typhi, adherence to intestinal epithelial cells mediated by fimbriae constitutes a key initial step in establishing infection. These adhesins can recognize specific receptors on host cell surfaces and/or extracellular matrix (ECM) components, including collagen, laminin, fibronectin, and heparan sulfate [1]. Laminin binding by various pilus and non-pilus bacterial adhesins represents an important initial step in the invasion process of numerous pathogenic microorganisms [2]. The adhesion of bacteria to the surface of the mucosa is an important factor in the early stages of the infection process [3, 4].
FimH is an adhesion protein located at the tip of type 1 fimbriae in S. Typhi, with a molecular weight of 36-kDa. Fimbriae are elongated protein appendages on the bacterial surface that facilitate host interaction, enhance environmental survival, aid in motility, promote colonization and cellular entry, and enable the exchange of genetic material [4-8]. The results of the in situ immuno-electron microscopy experiment by Hahn et al. [9] showed that the minor subunit (FimH) mediates the adhesion of the pilus to epithelial cells with a spring-like appearance. The FimG and FimF subunits connect FimH to the FimA rod; the orientation is sequential: FimA–FimF–FimG–FimH [9-12].
The fimbria, which are sticky hair-like structures, are made up of a cylindrical rod called pilus made from the FimA subunit, along with small tip fibers that include FimF, FimG, and FimH adhesions [13-15]. Fimbria type-1 is a component of fimbriae in S. Typhimurium, facilitating adherence and penetration of human epithelial cells and linked to phagocytosis by T cell hospes pediatrics-1 (THP-1) [16-18].
When bacteria reach the surface of the host cell, they will attach to it and undergo the colonization process [5, 19]. This occurrence is important, especially in the surface area of the mouth, small intestine, and bladder, as it is always washed with fluids [3, 20]. Only in this area are bacteria able to form adhesions to the mucosa, so they can stay and multiply [21, 22]. Two mechanisms for adhering to the host surface include pili or fimbrial adhesion and afimbrial adhesion (AFA) [23]. The newly identified mechanism involves bacteria secreting receptor proteins that target the interior of host cells, undergo phosphorylation, and bind to eukaryotic cell structures, thereby forming new receptors that enable stronger bacterial attachment [24-26]. On the other hand, the secretion of the protein hole receptor 90 (Hp90) in the previous host also provides another mechanism to inhibit bacterial adhesion and infection [27-30].
A model of the adhesion mechanism of potential bacteria was developed into an anti-adhesion vaccine to inhibit bacterial colonization and infection [31, 32]. The appearance of fimbria on the surface of bacteria is an antibody target to inhibit the adhesion or interaction of bacteria with the host [33, 34]. This adhesion receptor binding will activate the transduction of complex signals in host cells with a variety of consequences, including activation of innate host defenses and increased colonization and bacterial invasion [5].
Salmonella Typhi bacteria are a significant intracellular pathogen as they exclusively infect humans and induce typhoid fever. The S. Typhi bacterial infection is recognized as the etiological agent of enteric fever, presenting a complex clinical issue for physicians globally [1-3]. The use of E. coli, S. flexneri, and S. Typhi bacteria in this study aims to prove that the FimH36-kDa protein from S. Typhi has the same ability to inhibit the adhesion of the enterobacter bacterial group to enterocyte cells because it has similarities in receptor proteins and the function of FimH type-1 proteins [35-37]. Shigella flexneri and Enteropathogenic Escherichia coli (EPEC) are bacteria that cause moderate to severe diarrhea in young children in developing countries [38, 39]. Patients with mild infections caused by EPEC are not dehydrated, inflamed, fever, vomit, or experience abdominal pain [40-43].
Currently, several vaccines targeting Salmonella enterica serovar Typhi are licensed for human use. However, vaccine development is hindered by the antigenic diversity among the thousands of serovars capable of causing human infection. Conjugate vaccines, which consist of polysaccharides covalently linked to a protein, address this limitation. Although polysaccharides alone are poor immunogens, particularly in children, conjugation with a protein carrier enhances both humoral and cellular immune responses [41]. The currently available vaccines for Salmonella include the live-attenuated S. Typhi Ty21a and the purified Vi polysaccharide formulations. These vaccines demonstrate moderate efficacy (50–70%) in older children and adults but are not suitable for children under five years of age. In contrast, the recombinant exoprotein A (rEPA) Vi conjugate vaccine has shown promising results, achieving a protective efficacy of 91.1% [3]. Therefore, potential bacterial components can be used and developed as pure antigens for subunit or conjugate vaccines. This research is crucial for identifying biological materials that prevent the adherence of S. Typhi and other enterobacterial pathogens.
2.1 Research design
This research is exploratory and a laboratory experiment, with the research material being bacteria, Escherichia coli (O127:H6), Shigella flexneri (2457T), and Salmonella Typhi (endogenius). Agar Mac-Conkey (Oxoid), Thiaproline Carbonate Glutamat (TCG) Agar (Oxoid), BSA (Oxoid) and Heart Brain Infusion Broth (BHI), SSA (Oxoid), LB (Oxoid), TSIA(Oxoid), electrophoresis gel Sodium-dodecyl sulfate polyacrylamide (SDS-PAGE), and Balb/c mouse strain. Chemical reagent: TCA (Oxoid), ammonium sulfate, sodium chloride, sodium citrate, EDTA, EGT, dithiothreitol, KCl, Na2HPO4, KH2PO4, acrylamide, bis-acrylamide, glycine, tris-HCl, basa tris, SDS, Temed, ammonium perisulphate, coomassie blue, methanol PBS, glacial acetic acid, glycerol, bromothymol blue, 2-mercaptoethanol, Tween 20, p-nitrophenyl phosphate, diethanolamine, MgCl2. Gram staining, oxidase reagent, and protein marker (Wide Range) from Sigma Ultra [44-46].
Instruments used: spectrophotometer, micropipette, autoclave, incubator, colony counter, microscope, microtiter plate, dialysis membrane, electro-elution membrane, magnetic stirrer, centrifuge, Eppendorf 1.5 ml to 100 ml, elektroforesis vertikal (Bio-Red Mini Protein), blotting tool (Bio-Rad Trans-Blot SD-Semi-Dry Transfer Cell). Micropipette 5–20 μl, 50–200 μl, 200–1000 μl [47, 48].
2.2 Growth of bacteria
Strains of Escherichia coli, Shigella flexneri, and Salmonella Typhi were cultured on Moeller-Hinton Agar medium at a temperature of 37℃ for 18–24 hours [47]. Gram staining is performed on the growing bacterial colonies before inoculating them onto MacConkey differential media at 35℃ for 18-24 hours. S. Typhi colonies that grow on MacConkey media are re-cultured on selective BSA media at 37℃ for 18-24 hours [48, 49].
2.3 Isolation and fimbria protein separation from bacterial cells
Bacterial cultures grown in Mueller–Hinton Broth (MHB) were transferred into 100-cc centrifuge tubes. Trichloroacetic acid (TCA) was added to achieve a final concentration of 3%, followed by centrifugation at 6000 rpm for 15 min at 4℃. The resulting pellets were resuspended in phosphate-buffered saline (PBS; pH 7.4), and fimbriae were sheared using a modified Omni-mixer at 4℃. The fimbrial protein fraction was collected by centrifugation at 12,000 rpm for 15 min at 4℃. This shearing–centrifugation cycle was repeated until fimbrial fractions free of cellular debris were obtained. Protein isolation and purification via SDS-polyacrylamide gel electrophoresis (PAGE) was conducted with a mini-slab gel apparatus (Hoeffer Scientific Instruments, San Francisco, Calif.) by the method of Laemmli et al. [50]. Whole cells or protein fractions were solubilized in SDS sample buffer, stacked in 4.5% polyacrylamide (100 V), and separated in 12.5% polyacrylamide (200 V). Samples containing SEF 21 to be analyzed by SDS-PAGE required treatment at 100℃ for 5 min in SDS-PAGE sample buffer containing 0.2 M glycine (pH 7.4) before electrophoresis. Proteins were stained with Coomassie Brilliant Blue R-250 [48, 49]. Enterocyte cell models were isolated from intestinal enterocytes of Mus musculus Balb/c and Salmonella Typhi bacterial strains [8873].
2.4 Western blot analysis
Western blotting was performed according to Laemmli et al. [50] following SDS-PAGE of FimH protein. SDS-PAGE gels and nitrocellulose (NC) membranes were equilibrated in transfer buffer for 30 min and assembled in a semi-dry Trans-Blot system (Bio-Rad) in the order: filter paper–NC membrane–SDS-PAGE gel–filter paper. Protein transfer was conducted at 300 mA and 20 V for 2 h. Membranes were rinsed with distilled water, stained with Ponceau for 1 min, and destained with distilled water to verify transfer efficiency. Membranes were blocked with 5% skim milk in TBS overnight at 4℃ and washed with TBS-Tween 0.05% (1 × 10 min and 2×10 min, gentle shaking). The primary antibody (AdhO36-kDa, 1:100 in 1% skim milk/TBS) was incubated for 2 h at room temperature without shaking, followed by washing as above. Secondary antibody (biotin-conjugated anti-mouse IgG, 1:500 in TBS) was incubated for 1 h at room temperature without shaking. After washing (TBS-Tween 0.05%, 1 × 10 min, gentle shaking), membranes were incubated with SA-HRP for 40 min without shaking, followed by TMB-membrane and Western Blue AP substrates. The reaction was stopped with ddH₂O, membranes were air-dried, and scanned for documentation.
2.5 Dot blot assay
A Bio-Dot Apparatus (Bio-Rad) fitted with a pre-wetted nitrocellulose (NC) membrane (PBS, 5–10 min) was used for sample application. Fimbrial protein samples (50 μL in PBS containing 1% Na-azide) were loaded into each well and vacuumed until fully absorbed. The membrane was blocked with TBS containing 5% skim milk overnight at 4℃, then washed three times with TBS–Tween-20 (0.05%) for 3 min each. The membrane was incubated with primary antibody AdhO36 kDa (1:800 in TBS with 1% skim milk) for 60 min at room temperature, followed by two washes with TBS–Tween-20 (0.05%) for 10 min each. Biotin-conjugated anti-mouse IgG secondary antibody was applied for 60 min at room temperature, followed by the same washing procedure. SA–HRP (50 μL, 1:1000) was added and incubated for 40–60 min, after which the membrane was washed and developed with Western Blue substrate for ~20 min in the dark at room temperature. The reaction was stopped with distilled water, and the membrane was air-dried prior to scanning.
2.6 The biuret method is used to measure protein levels
Protein concentration was measured using the Biuret method. A total of 200 μL of fimbrial protein solution from Salmonella Typhi, purified by electroelution, was transferred into a microtube (Eppendorf) and mixed with 800 μL of Biuret reagent. The mixture was vortexed to ensure homogeneity and then incubated for 30 minutes. Absorbance was measured using a UV-VIS spectrophotometer at a wavelength range of 500–600 nm. A blank solution was prepared using 200 μL of distilled water and 800 μL of Biuret reagent.
2.7 Preparation for bacterial adhesion test
For the adhesion test, a group of bacterial cultures of Escherichia coli, Shigella flexneri, and Salmonella Typhi has been prepared. First, inoculate one loop of the bacterial colony into a test tube with 5 mL of nutrient broth, then incubate it for 24 hours at 37℃. Prepare a test tube with 5 mL of LB broth medium. Subsequently, introduce the bacterial culture into the LB medium, homogenize the mixture, and incubate for 24 hours at 37℃. Next, the bacterial culture is centrifuged at 6000 rpm for 15 minutes at 4℃. Take the suspended sediment in PBS with 1% BSA. Measure bacterial density (OD) using a spectrophotometer. The bacterial content is 108/ml (concentration at OD = 1, λ = 540 nm, for λ = 600 nm = 109/ml) [50, 51].
2.8 Mice’s enterocyte cell preparation and adhesion test adhesion
The results of the electroelution of the FimH36-kDa protein were prepared for adhesion tests on E. coli, S. flexneri, and S. Typhi. Doses of the FimH36-kDa protein were prepared at 0 μg (control), 100 μg, 200 μg, 300 μg, and 400 μg in 300 μl PBS. Each dose of fimbriae protein was added to 300 μl of enterocyte suspension and slowly homogenized in a water bath shaker at 37℃ for 30 minutes. Each dose of fimbrial protein was mixed with 300 μl of enterocyte suspension and gently stirred in a water bath shaker at 37℃ for 30 minutes. Next, this mixture is added to a bacterial suspension (108/mL), 300 μl. Incubate in a shaking incubator for 30 minutes at 37℃. Centrifuge at 1500 rpm at 4℃ for 3 minutes, then wash the sediment twice with PBS. Gather the precipitated sediment, apply it to a glass slide, and conduct Gram staining. The preparation was observed under a microscope with 100x magnification, and the number of bacteria adhering to the enterocytes was counted as the adhesion index. The adhesion index is the average number of bacteria adhering compared to the total number of bacteria available in an experiment per 100 enterocyte cells [52-54]. After obtaining the value of the bacterial adhesion index, the percentage reduction in the cell adhesion index was quantified. The negative control for mouse enterocytes was not treated with the 36-kDa FimH protein from S. Typhi (enterocytes mixed with bacterial cells).
3.1 Gram staining of E. coli, S. flexneri, and S. Typhi bacterial cells
To ensure the bacterial samples used in the research, a Gram stain was performed to observe cell morphology, and a Microbact system test was conducted to determine the biochemical reactions as part of the bacterial species identification. Here are the results of the bacterial Gram staining (Figure 1).
Figure 1. Gram staining of Gram-negative bacteria (A) Escherichia coli, (B) Salmonella Typhi, and (C) Shigella flexneri
The staining results were performed to confirm that the bacteria utilized in the research samples appropriately display the traits of Gram-negative bacteria. The identification results of the bacteria E. coli, S. flexneri, and S. Typhi accurately reflect their expected characteristics and are further validated through biochemical property tests. The biochemical experiments conducted with the Microbact system 2000 kit demonstrate that these three bacteria are capable of fermenting different glucose substrates. Consequently, these bacteria are suitable as test subjects for exposure to the FimH protein.
3.2 Results of the isolation of the FimH36-kDa protein of S. Typhi
Here are the results of the SDS-PAGE for the isolation of S. Typhi protein.
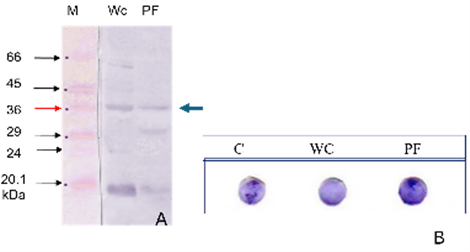
Figure 2. SDS-PAGE results of the FimH36-kDa protein from S. Typhi
Blood sample from a patient who has tested positive for typhoid fever from the Central Sulawesi Provincial Health Laboratory. The measured molecular weight of this protein is 36 kDa. M: Marker proteins, WC: Bacterial cell whole protein, PF: Fimbriae proteins, kDa: Kilo dalton. Several protein bands were detected, with molecular weights ranging from 20.1 kDa to 66 kDa. However, only the FimH protein with a molecular weight of 36 kDa was purified and utilized in this study.
The results of the study in Figure 2 show that the S. Typhi isolate from Palu has been proven to possess the FimH protein, as indicated by the results of the adhesin protein isolation. The FimH protein isolated from the endogenous S. Typhi of patients from the Central Sulawesi Health Laboratory was subsequently named FimH36-kDa due to its molecular weight of 36-kDa. Meanwhile, according to Zainer et al. [13], one unit of fimbriae consists of all the structural components of the proteins FimA, FimF, FimG, and FimH [55-57]. It is suspected that the FimH protein is much more susceptible to damage and depolymerization compared to the pure FimA protein fimbriae. Fimbriae type-1 are heteropolymer proteins because a single organelle contains about 1000 structural components of the FimA protein and, in addition, consists of three other proteins, FimF, FimG, and FimH, which make up 1–2% of the total fimbrial protein [12, 58, 59]. Likewise, Galán [23] and Sheikh et al. [60] explained that several molecular proteins ranging from 45 to 110 kDa are referred to as fimbriae type-1 receptors on different cells [35, 57, 58]. Referring to the low-range standard protein Sigma Marker M3913, 6.5–66 kDa, this protein is believed to be a subunit of the S. Typhi fimbriae protein.
Table 1. Description of immunofluorescence/confocal studies of FimH binding receptors
|
Types of Bacteria |
Description |
Reference |
|
E. coli |
Type 1 pili play an important role in optimal adhesion to the small intestinal epithelium. |
[60] |
|
E. coli |
The immunofluorescence microscopy image shows bacterial type 1 pili in the kidney (inset: green indicates anti–type 1 pili, and red indicates nuclear staining with SYTO 61). |
[61] |
|
E. coli |
The FimH type-1 fimbrial adhesin allows pathogenic Escherichia coli to adhere to glycoproteins in the epithelial linings of the human bladder and intestinal tract, by using multiple fimbriae simultaneously. |
[61] |
|
Salmonella |
Adhesion assay with ICE-PURO cells and ICE-CRT cells, Flow cytometry analysis of IPEC-CRT cells. |
[59] |
|
Salmonella |
A potential FimH receptor, endo-1,3-β-d-glucanase, has been identified, and the interaction was found to be strong and specific, with a dissociation constant in the nanomolar range. |
[62] |
|
Salmonella enterica typhi |
Analysis of bacterial binding upon surface labeling of S. enterica typhi Ty21a with FimH. |
[63] |
|
Shigella flexneri |
A representative spinning-disk confocal MIP of wild-type S. flexneri infection foci from 4 h postchallenge. Bacteria intrinsically express GFP (green); F-actin is phalloidin568 (red), and nuclei are DAPI-stained (blue). |
[64] |
The data in Table 1 are derived from previous studies that support the role and presence of the FimH protein as an adhesin factor capable of binding to receptors in the Enterobacter group, particularly in E. coli, S. Typhi, and S. flexneri. Several studies have revealed that FimH variants within the same Salmonella serotype may differ in their ability to recognize receptors in various tissue types, ultimately affecting their adhesion capability.
3.3 Western blot and dot blot results of FimH protein from S. Typhi
The Western blot assay was performed using the Towbin method [15] to determine whether the FimH protein could react with the polyclonal antibody AdhO36. Based on the qualitative observation of color gradients in Figure 3, the fimbrial adhesin protein was found to be recognized by the polyclonal antibody AdhO36. This finding is consistent with the characteristics of pilus proteins in Gram-negative bacteria, which function as mediators of adhesion and defense, enabling them to recognize and bind to at least one epitope molecule.
The presence of the FimH protein was detected through cross-reaction with the polyclonal antibody AdhO36 in a Dot blot assay, based on the presumed similarity in protein characteristics. The results demonstrated that the S. Typhi FimH protein reacted positively with the polyclonal antibody AdhO36, thereby confirming the presence of the target protein Figure 3.
Figure 3. Western blot and dot blot results of the FimH protein from S. Typhi
In part (A), M: molecular weight marker; WC: whole-cell proteins, PF: fimbrial proteins, kDa: kilodalton. In part (B), C: control whole-cell S. Typhi; WC: whole-cell proteins of S. Typhi; PF: fimbrial proteins of S. Typhi. In section (A), the Western blot results showed a consistent positive protein band at a molecular weight of 36 kDa, corresponding to the FimH36-kDa protein, along with additional bands at different molecular weights. The dot blot assay also yielded positive results, characterized by a specific reaction between the polyclonal antibody AdhO36 and the target FimH protein of S. Typhi. These findings confirm the presence of the target protein as intended in the study.
The concentration of FimH protein from S. Typhi, purified through electroelution and measured using the Biuret method, was determined to be 3.0125 µg/mL. These findings provide strong support for the entire isolation process and conclusively confirm the presence of the FimH protein in S. Typhi.
It is known that the FimA protein is the main subunit that forms 95% of the fimbrial shaft and is a structurally heterogeneous antigen among different species. The crystals of the FimH protein and the sugar-binding region have been mapped to the N-terminal half of the molecule (residues 1-156 aa), while the region associated with the fimbrial shaft has been mapped to the C-terminal half (residues 160-277 aa) of the FimH protein molecule [8]. Based on the research findings of Dufresne et al. [32], it is confirmed that the 20.1 kDa protein, which is the main component of the fimbrial shaft, can be reliably identified as the FimA protein of S. Typhi. Proteins with a molecular weight greater than 20.1 kDa are additional components, including the FimH protein of S. Typhi. Therefore, the expression of the S. Typhi FimH protein isolate can be trusted as a component of the FimH subunit protein because it has the same characteristics and adhesion capabilities as enterocytes [40, 61]. All these glycoprotein receptors have binding sites containing oligomannose common to the FimH protein, adhesion (mannose-binding subunit), found in fimbriae type-1 [8]. Therefore, the expressed proteins are a group of fimbrial proteins (fimbriae type-1 proteins), which act as adhesion media on host cells. Bacterial cells modify their physiology at many regulatory levels, including gene and protein expression, during adhesion or biofilm development [53, 59].
Until now, the crystal structure of FimH Salmonella Typhi is still in the process of being finalized, especially the design of structure-based inhibitors and the search for effective antagonist properties [57, 65-67]. The crystal structure of FimH from S. Typhi still requires a reliable and generally recognized homology modeling method. To adjust its three-dimensional structure, homology with the template structure of 1 klf of the Protein Data Bank (PDB) of E. coli FimH adhesion [6]. Although the crystal structure of FimH of S. Typhi has not been completely resolved, studies on other serovars suggest that allele variation in FimH can affect the adhesion capacity of bacteria [67, 68]. Small differences in amino acid sequences can have a significant impact on the ability of bacteria to attach to host cells [69].
Ligand-receptor interactions strengthened by mechanical stress, known as catch-bonds, play a key role in cell adhesion [70]. Pathogenic bacteria will adhere to the host epithelium through the adhesin FimH, a two-domain protein at the tip of type-1 pili that recognizes terminal mannose on epithelial glycoproteins. The result indicates the expression of the fimH gene associated with the FimH protein of S. Typhi. Similarly, this adhesion test proves that the FimH protein of S. Typhi is an adhesion protein, which is believed to be one of the virulence factors [56, 57].
3.4 Inhibition of FimH36-kDa protein adhesion on mouse enterocyte cells
The following shows the results of observing the adhesion of test bacteria to mouse enterocyte cells.
Figure 4. Adhesion of E. coli, S. Typhi, and S. flexneri cells to mouse enterocyte cells with concentrations of FimH36-kDa protein exposure (A=0 µl), (B=100 µl), (C=200 µl), (D=300 µl), and (E=400 µl)
In E. coli, S. Typhi, and S. flexneri, the letter A (control) denotes a mixture of bacteria and enterocyte cells without treatment with the FimH protein. Bacterial adhesion in the control group is noticeably higher compared to the treated groups. Letters B, C, D, and E correspond to increasing concentrations of FimH protein exposure, ranging from 100 µL to 400 µL. The arrows indicate bacterial adhesion sites, which decrease in number across all bacterial strains as the protein concentration increases. These findings suggest that the FimH protein plays a crucial role in inhibiting bacterial adhesion to enterocyte cells.
Microscopic observation results of the adhesion of E. coli, S. Typhi, and S. flexneri bacteria, Figure 4, to mouse enterocytes after exposure to FimH36-kDa protein at different concentrations. The observation of these bacterial cells was conducted through Gram staining and analyzed using a microscope with 100x magnification. The progressive decrease in the number of bacterial cells adhering to the number of bacterial cells is visible with increasing exposure to FimH36-kDa concentration. In the control group (A=0 µl), many bacteria were seen densely adhering to the surface of the enterocyte cells. This result indicates a high natural adhesion level of the bacteria to enterocytes without the intervention of the FimH protein.
The increased exposure to FimH36-kDa concentrations (B=100 µl to E=400 µl) shows a significant decrease in the number of bacteria that can adhere. At the highest concentration (E), adhesion is almost not visually detectable, indicating that the FimH protein works competitively to block mannosyl receptors on the surface of enterocyte cells that are usually used by bacterial fimbriae to attach. This means that FimH competes with other molecules (such as mannose) to bind to the receptor. The phenomenon of the decrease in the number of bacteria adhering appears to be consistent across the three bacterial species. This evidence indicates that their adhesion mechanisms likely utilize the same receptors [17] or similar ones, and that the FimH36-kDa protein of S. Typhi can disrupt cross-species adhesion of Gram-negative enteric pathogens [18].
3.5 Bacterial adhesion index exposed to FimH36-kDa protein S. Typhi
Based on the calculation of the adhesion index of E. coli, S. flexneri, and S. Typhi bacteria exposed to the FimH36-kDa S. Typhi protein, it can be seen in the following image.
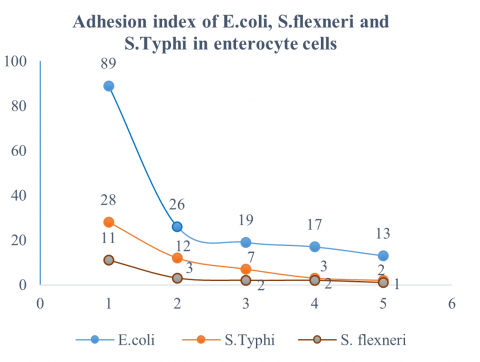
Figure 5. Adhesion index of E. coli, S. Typhi, and S. flexneri bacteria
Note: Protein FimH36-kDa concentration,1 = Control, 2 = 100 µl, 3 = 200 µl, 4 = 300 µl, and 5=400 µl
The calculation of the adhesion index of the FimH36-kDa protein against E. coli, S. flexneri, and S. Typhi has proven capable of reducing the number of bacterial adhesions to enterocyte cells. This can provide positive hope for becoming a vaccine candidate against the S. Typhi bacteria [58]. According to the results of this study, as shown in Figure 5, the FimH protein of S. Typhi is also capable of inhibiting the adhesion of E. coli, S. flexneri, and S. Typhi cells to mouse enterocyte cells. The reduction in the number of E. coli cell adhesions to mouse enterocytes indicates that this protein also has the potential to act as an anti-adhesion agent or vaccine component against these three bacteria, making them non-pathogenic to humans [56].
The exposure of the FimH36-kDa S. Typhi protein to enterocyte cell receptors reduces the binding sites and adhesion capability of the test bacteria's FimH protein. This occurs when there is a slight difference in the amino acid sequence. FimH protein can significantly affect the bacteria's ability to adhere to various types of cells [10, 71, 72], which in turn can affect virulence. The exposure of the FimH36-kDa protein in this study is believed to induce competitive binding with the fimbriae of pathogenic bacteria, thereby preventing bacterial adherence to enterocytes. This mechanism is explained through the concept of competition, where soluble FimH attaches to the mannose receptor on the cell surface, blocking potential binding sites for the fimbriae of the bacteria S. Typhi, S. flexneri, and E. coli. As a result, the bacteria lose their ability to adhere optimally, which in turn reduces the initial colonization rate in the intestinal mucosa. This change is very relevant, considering that bacterial adhesion is a crucial stage in the infection process, facilitating colonization, invasion, and activation of the host's immune response.
Inhibition of the adhesion of E. coli, S. flexneri, and S. Typhi bacteria by the FimH36-kDa protein of S. Typhi is expected to be a new study on developing vaccines for human Gram-negative bacterial pathogens [73]. This is supported by research findings that show that the higher the concentration of exposure to the FimH36-kDa S. Typhi protein, the better its adhesion inhibition capacity [32]. The reduction in adhesion index is as follows: E. coli decreased from 89 to 13 cells, S. Typhi from 28 to 2 cells, and S. flexneri from 11 to 1 cell per enterocyte.
The following table displays the results of computing the percentage reduction in the bacterial adhesion index.
Table 2. Percentage reduction in the number of bacterial cell adhesion index in enterocyte cells
|
FimH36-kDa Protein Treatment |
Reduction in the Number of Bacterial Adhesion Indexes (%) |
||
|
E. coli |
S. Typhi |
S. flexneri |
|
|
K |
0 |
0 |
0 |
|
A |
70.79 |
57.15 |
72.73 |
|
B |
78.66 |
75.01 |
81.80 |
|
C |
80.9 |
89.30 |
81.80 |
|
D |
85.4 |
92.86 |
90.91 |
Note: Protein FimH36-kDa concentration, K = Control, A = 100 µL, B = 200 µL, C = 300 µL, and D=400 µL
Table 2 shows the percentage reduction in adhesion index for three bacterial strains. In the negative control (K), enterocytes were exposed only to bacteria, while treatments A–D received increasing concentrations of FimH protein (100–400 µL). Higher FimH concentrations corresponded to greater reductions in adhesion.
FimH36-kDa protein treatment elicited a pronounced, dose-dependent decrease in the adhesion index of E. coli, S. Typhi, and S. flexneri, with the greatest reduction observed at 400 µL (85.4%, 92.86%, and 90.91%, respectively). S. Typhi demonstrated the highest sensitivity, particularly at higher protein concentrations. The FimH protein uses its carbohydrate-binding domain (CBD) to recognize mannose residues on cell surface receptors. The strength of the bond is highly dependent on specific oligomannoside structures, such as Manα1-3Manβ1-4GlcNAc, which is an optimal target for FimH with a high affinity (kₙ ≈ 20 nM) for PMC +1 [74].
The FimH36-kDa protein, structurally analogous to the FimH domain of Salmonella Typhi, acts as a potent competitive antagonist by preferentially binding to mannosylated receptors with higher affinity than native fimbriae, thereby preventing bacterial adhesion to host cells [32]. The high adhesion effectiveness of FimH36-kDa to S. Typhi is attributable to its strong affinity for mannosylated receptors, structural similarity, and specific molecular interactions, which collectively enhance its competitive binding ability against bacterial adhesion, given that both originate from the same species.
This reduction presents definitive proof that the FimH36-kDa protein from S. Typhi demonstrates significant anti-adhesive effects against enteric pathogens in vitro. FimH36-kDa S. Typhi protein, which acts as a virulence factor and antiadhesive medium [5, 23, 65], is expected to inhibit the initial stage of bacterial adhesion activity and can be further studied as a vaccine candidate against S. Typhi bacteria and other enterobacterial groups. Although the results of this study are promising, further research is needed on the effectiveness of the more complex FimH36-kDa protein, such as testing at various concentrations, exposure durations, toxicity tests, and host immunological responses, before clinical application can be recommended.
This study concludes that the FimH36-kDa protein from the Palu isolate of S. Typhi effectively inhibits the adhesion of Escherichia coli, Shigella flexneri, and Salmonella Typhi, evidenced by a significant reduction in the bacterial adhesion index on mouse enterocytes following exposure to the FimH36-kDa protein. This finding is a biological element that suggests its potential as a candidate for a typhoid vaccine to prevent colonization and infection by the pathogen Enterobacter. Despite the encouraging results, additional research is required to assess the efficacy of the FimH36-kDa protein. The limitation of this study is the absence of immunogenicity testing to evaluate the ability of the FimH36-kDa protein antigen to induce antibody production, as well as its capacity to trigger innate and adaptive immune responses, either in vitro or in vivo.
I sincerely extend my gratitude for the support and funding provided through the Faculty of Teacher Training and Education Grant at Tadulako University for the 2023 fiscal year, under Contract No.: 6805/UN28/KU/2023.
[1] Pierrat, X., Wong, J.P., Al-Mayyah, Z., Persat, A. (2021). The mammalian membrane microenvironment regulates the sequential attachment of bacteria to host cells, MBio. 12(4): 1-16. https://doi.org/10.1128/mBio.01392-21
[2] Schéele, S., Nyström, A., Durbeej, M., Talts, J.F., Ekblom, M., Ekblom, P. (2007). Laminin isoforms in development and disease. Journal of Molecular Medicine, 85(8): 825-836. https://doi.org/10.1007/s00109-007-0182-5
[3] Ghosh, S., Chakraborty, K., Nagaraja, T., Basak, S., Koley, H., et al. (2011). An adhesion protein of Salmonella enterica serovar Typhi is required for pathogenesis and potential target for vaccine development. Proceedings of the National Academy of Sciences, 108(8): 3348-3353. https://doi.org/10.1073/pnas.1016180108
[4] Li, H., Xu, D., Tan, X., Huang, D., Huang, Y., et al. (2023). The role of trehalose biosynthesis on mycolate composition and L-glutamate production in Corynebacterium glutamicum. Microbiological Research, 267: 127260. https://doi.org/10.1016/j.micres.2022.127260
[5] Darmawati, S., Ethica, S.N., Prastiyanto, M.E., Depamede, S.N., Putri, E.O., Kamaruddin, M. (2022). Molecular characterization of a 42 kDa subunit pili protein of Salmonella typhi causes typhoid fever. Biodiversitas Journal of Biological Diversity, 23(2): 962-968. https://doi.org/10.13057/biodiv/d230239
[6] Kang, X., Chen, J., Zhou, X., Ed-Dra, A., Yue, M. (2023). Allelic functional variation of FimH among Salmonella enterica subspecies. Animal Diseases, 3(1): 33. https://doi.org/10.1186/s44149-023-00101-y
[7] Azam, M.W., Zarrilli, R., Khan, A.U. (2023). Updates on the virulence factors produced by multidrug-resistant Enterobacterales and strategies to control their infections. Microorganisms, 11(8): 1901. https://doi.org/10.3390/microorganisms11081901
[8] Gahlot, D.K., Taheri, N., MacIntyre, S. (2022). Diversity in genetic regulation of bacterial fimbriae assembled by the chaperone usher pathway. International Journal of Molecular Sciences, 24(1): 161. https://doi.org/10.3390/ijms24010161
[9] Hahn, E., Wild, P., Hermanns, U., Sebbel, P., Glockshuber, R., et al. (2002). Exploring the 3D molecular architecture of Escherichia coli type 1 pili. Journal of Molecular Biology, 323(5): 845-857. https://doi.org/10.1016/S0022-2836(02)01005-7
[10] Uchiya, K.I., Kamimura, Y., Jusakon, A., Nikai, T. (2019). Salmonella fimbrial protein FimH is involved in expression of proinflammatory cytokines in a toll-like receptor 4-dependent manner. Infection and Immunity, 87(3): 10-1128. https://doi.org/10.1128/IAI.00881-18
[11] Baxter, M.A., Jones, B.D. (2015). Two-component regulators control hilA expression by controlling fimZ and hilE expression within Salmonella enterica serovar Typhimurium. Infection and Immunity, 83(3): 978-985. https://doi.org/10.1128/IAI.02506-14
[12] Rehman, T., Yin, L., Latif, M.B., Chen, J., Wang, K., et al. (2019). Adhesive mechanism of different Salmonella fimbrial adhesins. Microbial Pathogenesis, 137: 103748. https://doi.org/10.1016/j.micpath.2019.103748
[13] Zeiner, S.A., Dwyer, B.E., Clegg, S. (2012). FimA, FimF, and FimH are necessary for assembly of type 1 fimbriae on Salmonella enterica serovar Typhimurium. Infection and Immunity, 80(9): 3289-3296. https://doi.org/10.1128/IAI.00331-12
[14] Bessaiah, H., Anamalé, C., Sung, J., Dozois, C.M. (2021). What flips the switch? Signals and stress regulating extraintestinal pathogenic Escherichia coli type 1 fimbriae (pili). Microorganisms, 10(1): 5. https://doi.org/10.3390/microorganisms10010005
[15] Schultz, B.M., Melo-Gonzalez, F., Salazar, G.A., Porto, B.N., Riedel, C.A., Kalergis, A.M., Bueno, S.M. (2021). New insights on the early interaction between typhoid and non-typhoid Salmonella serovars and the host cells. Frontiers in Microbiology, 12: 647044. https://doi.org/10.3389/fmicb.2021.647044
[16] Nasi, G.I., Georgakopoulou, K.I., Theodoropoulou, M.K., Papandreou, N.C., Chrysina, E.D., Tsiolaki, P.L., Iconomidou, V.A. (2023). Bacterial lectin FimH and its aggregation hot-spots: An alternative strategy against uropathogenic Escherichia coli. Pharmaceutics, 15(3): 1018. https://doi.org/10.3390/pharmaceutics15031018
[17] Frutos-Grilo, E., Kreling, V., Hensel, A., Campoy, S. (2023). Host-pathogen interaction: Enterobacter cloacae exerts different adhesion and invasion capacities against different host cell types. PloS One, 18(10): e0289334. https://doi.org/10.1371/journal.pone.0289334
[18] Fattinger, S.A., Sellin, M.E., Hardt, W.D. (2021). Salmonella effector driven invasion of the gut epithelium: Breaking in and setting the house on fire. Current Opinion in Microbiology, 64: 9-18. https://doi.org/10.1016/j.mib.2021.08.007
[19] Castello, A., Piraino, C., Butera, G., Alio, V., Cardamone, C., et al. (2023). Prevalence and antimicrobial resistance profiles of Salmonella spp. in poultry meat. Italian Journal of Food Safety, 12(2): 11135. https://doi.org/10.4081/ijfs.2023.11135
[20] Van Eyssen, S.R., Samarkina, A., Isbilen, O., Zeden, M.S., Volkan, E. (2023). FimH and Type 1 Pili mediated tumor cell cytotoxicity by uropathogenic Escherichia coli in vitro. Pathogens, 12(6): 751. https://doi.org/10.3390/pathogens12060751
[21] Li, Q. (2022). Mechanisms for the invasion and dissemination of Salmonella. Canadian Journal of Infectious Diseases and Medical Microbiology, 2022(1): 2655801. https://doi.org/10.1155/2022/2655801
[22] Whelan, S., Lucey, B., Finn, K. (2023). Uropathogenic Escherichia coli (UPEC)-associated urinary tract infections: The molecular basis for challenges to effective treatment. Microorganisms, 11(9): 2169. https://doi.org/10.3390/microorganisms11092169
[23] Galán, J.E. (2022). Salmonella Typhimurium and inflammation: A pathogen-centric affair. Nature Reviews Microbiology, 19(11): 716-725. https://doi.org/10.1038/s41579-021-00561-4
[24] Bao, H., Wang, S., Zhao, J.H., Liu, S.L. (2020). Salmonella secretion systems: Differential roles in pathogen-host interactions. Microbiological Research, 241: 126591. https://doi.org/10.1016/j.micres.2020.126591
[25] Ge, J., Wang, Y., Li, X., Lu, Q., Yu, H., et al. (2024). Phosphorylation of caspases by a bacterial kinase inhibits host programmed cell death. Nature Communications, 15(1): 8464. https://doi.org/10.1038/s41467-024-52817-1
[26] Kisiela, D.I., Chattopadhyay, S., Libby, S.J., Karlinsey, J.E., Fang, F.C., et al. (2012). Evolution of Salmonella enterica virulence via point mutations in the fimbrial adhesin. PLoS Pathogens, 8(6): e1002733. https://doi.org/10.1371/journal.ppat.1002733
[27] Feng, Y., Wang, S., Liu, X., Han, Y., Xu, H., et al. (2023). Geometric constraint-triggered collagen expression mediates bacterial-host adhesion. Nature Communications, 14(1): 8165. https://doi.org/10.1038/s41467-023-43827-6
[28] Chen, H., Schifferli, D.M. (2003). Construction, characterization, and immunogenicity of an attenuated Salmonella enterica serovar typhimurium pgtE vaccine expressing fimbriae with integrated viral epitopes from the spiC promoter. Infection and Immunity, 71(8): 4664-4673. https://doi.org/10.1128/IAI.71.8.4664-4673.2003
[29] Yue, M., Schifferli, D.M. (2014). Allelic variation in Salmonella: An underappreciated driver of adaptation and virulence. Frontiers in Microbiology, 4: 419. https://doi.org/10.3389/fmicb.2013.00419
[30] Zhou, G., Zhao, Y., Ma, Q., Li, Q., Wang, S., Shi, H. (2023). Manipulation of host immune defenses by effector proteins delivered from multiple secretion systems of Salmonella and its application in vaccine research. Frontiers in Immunology, 14: 1152017. https://doi.org/10.3389/fimmu.2023.1152017
[31] Sharma, A., Garg, A., Ramana, J., Gupta, D. (2023). VirulentPred 2.0: An improved method for prediction of virulent proteins in bacterial pathogens. Protein Science, 32(12): e4808. https://doi.org/10.1002/pro.4808
[32] Dufresne, K., Saulnier-Bellemare, J., Daigle, F. (2018). Functional analysis of the chaperone-usher fimbrial gene clusters of Salmonella enterica serovar Typhi. Frontiers in Cellular and Infection Microbiology, 8: 26. https://doi.org/10.3389/fcimb.2018.00026
[33] Gadar, K., McCarthy, R.R. (2023). Using next generation antimicrobials to target the mechanisms of infection. npj Antimicrobials and Resistance, 1(1): 11. https://doi.org/10.1038/s44259-023-00011-6
[34] Ledeboer, N.A., Frye, J.G., McClelland, M., Jones, B.D. (2006). Salmonella enterica serovar Typhimurium requires the Lpf, Pef, and Tafi fimbriae for biofilm formation on HEp-2 tissue culture cells and chicken intestinal epithelium. Infection and Immunity, 74(6): 3156-3169. https://doi.org/10.1128/IAI.01428-05
[35] Saha, T., Arisoyin, A. E., Bollu, B., Ashok, T., Babu, A., et al. (2023). Enteric fever: Diagnostic challenges and the importance of early intervention. Cureus, 15(7): 153589. https://doi.org/10.7759/cureus.41831
[36] Khanam, F., Ross, A.G., McMillan, N.A., Qadri, F. (2022). Toward typhoid fever elimination. International Journal of Infectious Diseases, 119: 41-43. https://doi.org/10.1016/j.ijid.2022.03.036
[37] Isidro-Coxca, M.I., Ortiz-Jiménez, S., Puente, J.L. (2024). Type 1 fimbria and P pili: Regulatory mechanisms of the prototypical members of the chaperone-usher fimbrial family. Archives of Microbiology, 206(9): 373. https://doi.org/10.1007/s00203-024-04092-3
[38] Ledwaba, S.E., Bolick, D.T., de Medeiros, P.H.Q.S., Kolling, G.L., Traore, A.N., et al. (2022). Enteropathogenic Escherichia coli (EPEC) expressing a non-functional bundle-forming pili (BFP) also leads to increased growth failure and intestinal inflammation in C57BL/6 mice. Brazilian Journal of Microbiology, 53(4): 1781-1787. https://doi.org/10.1007/s42770-022-00802-5
[39] Mare, A.D., Ciurea, C.N., Man, A., Tudor, B., Moldovan, V., Decean, L., Toma, F. (2021). Enteropathogenic Escherichia coli—A summary of the literature. Gastroenterology Insights, 12(1): 28-40. https://doi.org/10.3390/Gastroent12010004
[40] Kaur, P., Dudeja, P.K. (2023). Pathophysiology of enteropathogenic Escherichia coli-induced diarrhea. Newborn (Clarksville, Md.), 2(1): 102. https://doi.org/10.5005/jp-journals-11002-0056
[41] Rappuoli, R. (2018). Glycoconjugate vaccines: Principles and mechanisms. Science Translational Medicine, 10(456): eaat4615. https://doi.org/10.1126/scitranslmed.aat4615
[42] Hernández-Lucas, I., Gallego-Hernández, A.L., Encarnación, S., Fernández-Mora, M., Martínez-Batallar, A.G., et al. (2008). The LysR-type transcriptional regulator LeuO controls expression of several genes in Salmonella enterica serovar Typhi. Journal of Bacteriology, 190(5): 1658-1670. https://doi.org/10.1128/JB.01649-07
[43] Rabbani, S., Fiege, B., Eris, D., Silbermann, M., Jakob, R.P., et al. (2018). Conformational switch of the bacterial adhesin FimH in the absence of the regulatory domain: Engineering a minimalistic allosteric system. Journal of Biological Chemistry, 293(5): 1835-1849. https://doi.org/10.1074/jbc.M117.802942
[44] Simoens, L., Fijalkowski, I., Van Damme, P. (2023). Exposing the small protein load of bacterial life. FEMS Microbiology Reviews, 47(6): fuad063. https://doi.org/10.1093/femsre/fuad063
[45] Patel, S., Mathivanan, N., Goyal, A. (2017). Bacterial adhesins, the pathogenic weapons to trick host defense arsenal. Biomedicine & Pharmacotherapy, 93: 763-771. https://doi.org/10.1016/j.biopha.2017.06.102
[46] Melamed, S., Zaffryar-Eilot, S., Nadjar-Boger, E., Aviram, R., Zhao, H., et al. (2023). Initiation of fibronectin fibrillogenesis is an enzyme-dependent process. Cell Reports, 42(5): 112473, https://doi.org/10.1016/j.celrep.2023.112473
[47] Rodríguez-González, S., González-Dávalos, L., Robles-Rodríguez, C., Lozano-Flores, C., Varela-Echavarría, A., Shimada, A., Mora-Izaguirre, O. (2023). Isolation of bacterial consortia with probiotic potential from the rumen of tropical calves. Journal of Animal Physiology and Animal Nutrition, 107(1): 62-76. https://doi.org/10.1111/jpn.13699
[48] Hamada, N., Sojar, H.T., Cho, M.I., Genco, R.J. (1996). Isolation and characterization of a minor fimbria from Porphyromonas gingivalis. Infection and Immunity, 64(11): 4788-4794. https://doi.org/10.1128/iai.64.11.4788-4794.1996
[49] Gibson, D.L., White, A.P., Rajotte, C.M., Kay, W.W. (2007). AgfC and AgfE facilitate extracellular thin aggregative fimbriae synthesis in Salmonella enteritidis. Microbiology, 153(4): 1131-1140. https://doi.org/10.1099/mic.0.2006/000935-0
[50] Laemmli, U.K., Rasool, S.I., De Bergh, C. (1970). Nature Publishing Group. 228: 726-734. http://www.mendeley.com/research/discreteness-conductance-chnge-n-bimolecular-lipid-membrane-presence-certin-antibiotics/.
[51] Chagnot, C., Zorgani, M.A., Astruc, T., Desvaux, M. (2013). Proteinaceous determinants of surface colonization in bacteria: Bacterial adhesion and biofilm formation from a protein secretion perspective. Frontiers in Microbiology, 4: 303. https://doi.org/10.3389/fmicb.2013.00303
[52] Ifedinezi, O.V., Nnaji, N.D., Anumudu, C.K., Ekwueme, C.T., Uhegwu, C.C., et al. (2024). Environmental antimicrobial resistance: Implications for food safety and public health. Antibiotics, 13(11): 1087. https://doi.org/10.3390/antibiotics13111087
[53] Vakili, S., Haeili, M., Feizi, A., Moghaddasi, K., Omrani, M., Ghodousi, A., Cirillo, D.M. (2025). Whole-genome sequencing-based characterization of Salmonella enterica Serovar Enteritidis and Kentucky isolated from laying hens in northwest of Iran, 2022–2023. Gut Pathogens, 17(1): 2. https://doi.org/10.1186/s13099-025-00679-3
[54] Tempera, G., Corsello, S., Genovese, C., Caruso, F.E., Nicolosi, D. (2010). Inhibitory activity of cranberry extract on the bacterial adhesiveness in the urine of women: An ex-vivo study. International Journal of Immunopathology and Pharmacology, 23(2): 611-618. https://doi.org/10.1177/039463201002300223
[55] Sarowska, J., Futoma-Koloch, B., Jama-Kmiecik, A., Frej-Madrzak, M., Ksiazczyk, M., Bugla-Ploskonska, G., Choroszy-Krol, I. (2019). Virulence factors, prevalence and potential transmission of extraintestinal pathogenic Escherichia coli isolated from different sources: Recent reports. Gut Pathogens, 11(1): 10. https://doi.org/10.1186/s13099-019-0290-0
[56] Giese, C., Puorger, C., Ignatov, O., Bečárová, Z., Weber, M.E., et al. (2023). Stochastic chain termination in bacterial pilus assembly. Nature Communications, 14(1): 7718. https://doi.org/10.1038/s41467-023-43449-y
[57] Baxter, M.A., Jones, B.D. (2005). The fimYZ genes regulate Salmonella enterica serovar Typhimurium invasion in addition to type 1 fimbrial expression and bacterial motility. Infection and Immunity, 73(3): 1377-1385. https://doi.org/10.1128/IAI.73.3.1377-1385.2005
[58] Bager, A.M., Naeem, A.K. (2022). Genotypic detection of LifA and FimH genes in enter-pathogenic E. coli isolated from children suffering from diarrhea in AL-Najaf City, Iraq. International Journal of Health Sciences, 5: 1314-1321. https://doi.org/10.53730/ijhs.v6ns5.8865
[59] Grzymajlo, K., Ugorski, M., Suchanski, J., Kedzierska, A.E., Kolenda, R., et al. (2017). The novel type 1 fimbriae FimH receptor calreticulin plays a role in Salmonella host specificity. Frontiers in Cellular and Infection Microbiology, 7: 326. https://doi.org/10.3389/fcimb.2017.00326
[60] Sheikh, A., Rashu, R., Begum, Y.A., Kuhlman, F.M., Ciorba, M.A., et al. (2017). Highly conserved type 1 pili promote enterotoxigenic E. coli pathogen-host interactions. PLoS Neglected Tropical Diseases, 11(5): e0005586. https://doi.org/10.1371/journal.pntd.0005586
[61] McLellan, L.K., McAllaster, M.R., Kim, A.S., Tóthová, Ľ., Olson, P.D., et al. (2021). A host receptor enables type 1 pilus-mediated pathogenesis of Escherichia coli pyelonephritis. PLoS Pathogens, 17(1): e1009314. https://doi.org/10.1371/Journal.Ppat.1009314
[62] Waszczuk, W., Czajkowska, J., Dutkiewicz, A., Klasa, B., Carolak, E., Aleksandrowicz, A., Grzymajlo, K. (2024). It takes two to attach-endo-1, 3-β-d-glucanase as a potential receptor of mannose-independent, FimH-dependent Salmonella Typhimurium binding to spinach leaves. Food Microbiology, 121: 104519. https://doi.org/10.1016/j.fm.2024.104519
[63] Burggraaf, M.J., Waanders, L., Verlaan, M., Maaskant, J., Houben, D., et al. (2021). Surface labeling with adhesion protein FimH improves binding of immunotherapeutic agent salmonella Ty21a to the bladder epithelium. Bladder Cancer, 7(1): 79-90. https://doi.org/10.3233/BLC-200382
[64] Arena, E.T., Campbell-Valois, F.X., Tinevez, J.Y., Nigro, G., Sachse, M., et al. (2015). Bioimage analysis of Shigella infection reveals targeting of colonic crypts. Proceedings of the National Academy of Sciences, 112(25): E3282-E3290. https://doi.org/10.1073/pnas.1509091112
[65] Walker, W.H., Sanborn, B.M., Habener, J.F. (1994). An isoform of transcription factor CREM expressed during spermatogenesis lacks the phosphorylation domain and represses cAMP-induced transcription. Proceedings of the National Academy of Sciences, 91(26): 12423-12427. https://doi.org/10.1073/pnas.91.26.12423
[66] Ribet, D., Cossart, P. (2015). How bacterial pathogens colonize their hosts and invade deeper tissues. Microbes and Infection, 17(3): 173-183. https://doi.org/10.1016/j.micinf.2015.01.004
[67] Kolenda, R., Ugorski, M., Grzymajlo, K. (2019). Everything you always wanted to know about Salmonella type 1 fimbriae, but were afraid to ask. Frontiers in Microbiology, 10: 1017. https://doi.org/10.3389/fmicb.2019.01017
[68] Soto, G.E., Hultgren, S.J. (1999). Bacterial adhesins: Common themes and variations in architecture and assembly. Journal of Bacteriology, 181(4): 1059-1071. https://doi.org/10.1128/jb.181.4.1059-1071.1999
[69] Baker, E.P., Sayegh, R., Kohler, K.M., Borman, W., Goodfellow, C.K., Brush, E.R., Barber, M.F. (2022). Evolution of host-microbe cell adherence by receptor domain shuffling. Elife, 11: e73330. https://doi.org/10.7554/eLife.73330
[70] Sicard, J.F., Le Bihan, G., Vogeleer, P., Jacques, M., Harel, J. (2017). Interactions of intestinal bacteria with components of the intestinal mucus. Frontiers in Cellular and Infection Microbiology, 7: 387. https://doi.org/10.3389/fcimb.2017.00387
[71] Werneburg, G.T., Thanassi, D.G. (2018). Pili assembled by the chaperone/usher pathway in Escherichia coli and Salmonella. EcoSal Plus, 8(1): 10-1128. https://doi.org/10.1128/ecosalplus.esp-0007-2017
[72] Widyastuti, H., Santoso, S., Winarsih, S., Poeranto, S. (2023). The immunological response of Salmonella typhi protein subunits (Outer membrane protein 36 kDa, fimbriae 36 kDa, and flagellin) as a candidate for oral typhoid vaccine. Research Journal of Pharmacy and Technology, 16(8): 3891-3898. https://doi.org/10.52711/0974-360X.2023.00641
[73] Chevalier, G., Laveissière, A., Desachy, G., Barnich, N., Sivignon, A., et al. (2021). Blockage of bacterial FimH prevents mucosal inflammation associated with Crohn’s disease. Microbiome, 9(1): 176. https://doi.org/10.1186/s40168-021-01135-5
[74] Bouckaert, J., Mackenzie, J., De Paz, J.L., Chipwaza, B., Choudhury, D., et al. (2006). The affinity of the FimH fimbrial adhesin is receptor-driven and quasi-independent of Escherichia coli pathotypes. Molecular Microbiology, 61(6): 1556-1568. https://doi.org/10.1111/j.1365-2958.2006.05352.x