Sumardi![]() | Sapto Indrioko*
| Sapto Indrioko*![]() | Yeni Widyana Nurchahyani Ratnaningrum
| Yeni Widyana Nurchahyani Ratnaningrum![]() | Ida Luh Gede Nurtjahjaningsih
| Ida Luh Gede Nurtjahjaningsih![]()
© 2025 The authors. This article is published by IIETA and is licensed under the CC BY 4.0 license (http://creativecommons.org/licenses/by/4.0/).
OPEN ACCESS
The Seed Production Area (SPA) of sandalwood (Santalum album L.) in East Nusa Tenggara is Indonesia's sole designated site for sandalwood seed production. It was established 32 years ago using trees from six provenances on Timor Island. The SPA supports regional economies and ecological protection by producing high-quality seeds for commercially valuable trees and species conservation. Despite annual seed collection, information on reproductive status, genetic diversity, and genetic drift is limited. This study assessed fertility variation, genetic diversity, and the impacts of genetic drift on sandalwood seed production. Fertility was assessed through flower and fruit production, while genetic diversity was calculated using fertility and genetic drift data. The results showed high fertility variation (2.98), high genetic diversity (0.97), and low co-ancestry (0.03). This study revealed that 14.04% of trees contributed over 50% of the reproductive output. The effective population size (19.15%) indicates that a small percentage of trees contribute highly to seed production. To avoid dominance by highly fertile trees, seed collection is limited and mixed equally. These results provide a scientific basis for strategic seed production and genetic resource management. Recommendations include selectively pruning the crowns of highly fertile trees, increasing the number of fertile trees, and stimulating less fertile individuals to increase seed production while maintaining genetic diversity in the SPA.
Santalum album, seed production area, fertility, co-ancestry, effective population size
Sandalwood (Santalum album Linn.) is a highly valuable plant due to the santalol content of the essential oil produced from its heartwood, which is used in incense, perfume, medicine, and crafts [1-4]. This species is classified under the genus Santalum and is naturally found in the South Pacific Islands, Papua New Guinea, Indonesia, India, and Australia [5-7]. It faces a significant population decline due to overharvesting, driven by its economic value. Additional threats, including exploitation, deforestation, wildfires, low planting success, socioeconomic conditions, ownership status, grazing, and pest and disease infestations, exacerbate this decline [8, 9]. As a consequence, sandalwood is categorized as vulnerable by the IUCN [10]. The decline of sandalwood populations is currently a serious concern in Indonesia and worldwide. Other factors, such as low seed production, inconsistent germination, and poor seedling uniformity, further hinder its cultivation. Meanwhile, seedlings remain the primary method of propagation in regions like Indonesia, India, and Australia [11].
Research on sandalwood's reproductive traits reveals a self-incompatibility mechanism, indicating that this species tends to favor outcrossing [12-14]. In Western Australia, mating incompatibility has been observed in S. album, S. lanceolatum, and S. spicatum due to differences in their floral sexual organs [14]. Sandalwood flowers display protandrous dichogamy, where male organs mature before female ones, with pollen maturing at anthesis and pistils becoming receptive later as pollen viability decreases [15]. This asynchrony, along with non-panmixia (the inability of random pollen to fertilize pistils), can result in pollination failure [16]. Disproportionate contributions from gametes to the seed source can influence the genetic composition of the offspring and lead to reduced genetic diversity [17]. Their floral phenology influences the exchange of genes among trees and the genetic makeup of their offspring [18]. Achieving genetic excellence in the seed source necessitates complete phenological synchronization among the parents [19]. Lack of synchronization between female and male elders reduces the effective population size, leading to an imbalance in contributions from parent trees, an imbalance in self-fertilization rates, and an increased risk of contamination in the seed source [19, 20]. Research from numerous studies in seed sources indicates that asynchrony is a serious problem in seed production [21].
Sandalwood's high outcrossing nature makes genetic diversity essential for adequate fruit and seed production [12-14, 22]. Therefore, low selection intensity is used in breeding strategies to preserve this variety in tropical regions where the SPA serves as the primary seed source [23]. This is because high-intensity selection can cause unequal genetic contributions from male and female flowers, leading to genetic drift and reduced seed quality [23]. This can hamper conservation and breeding programs for sandalwood with low fruit production traits. Therefore, seed collection should be based on flower abundance and fertility to minimize genetic drift caused by flowering imbalances [17].
It is important to look at fertility when managing genetic resources and conservation areas. Fertility as a predictor has many benefits, including being simple to find, inexpensive, and useful in many areas of forest management. Fertility as an indicator has several advantages, such as being easily accessible, cost-effective, and useful in many areas of forest management [24]. The objectives encompass the production of high-quality seed, the development of tree breeding programs, and the design of appropriate seed collection systems [22, 25-27]. Moreover, fertility data offers directions to managers for ensuring sustained seed production and preserving the genetic diversity of trees [28]. Fertility is assessed using several factors associated with the reproductive structures of trees, including flowers and fruits [29-31]. Related studies have investigated fertility data derived from the strobili, seeds, cones, and fruits of both natural and artificial populations of several forest tree species [21-23, 25, 27, 30, 32-36]. The data presented in those studies not only elucidate the reproductive potential of species but also aid in understanding population dynamics and genetic adaptability. Moreover, they also provide theoretical approaches to understanding the biological mechanisms of fertility, as well as applied approaches focused on improving forest productivity, protecting genetic diversity, and adapting to environmental change. These research studies claim that an enhanced understanding of fertility can facilitate the implementation of strategic measures to optimize genetic resource management for future conservation and breeding purposes. The sandalwood seed production area (SPA) in East Nusa Tenggara (ENT), Indonesia's sole SPA, is the key source of superior genetic material. This species flowers twice a year in ENT, with peak seasons in June and December; the peak flower season is more in December than in June [37]. Seed requirements for planting in ENT were largely met with random seeds due to the inadequate seed production from sandalwood SPAs. Despite annual seed collection from the sandalwood SPA in ENT, data on fertility, genetic diversity, and potential genetic drift in future generations are lacking. Consequently, it is crucial to understand several factors that are essential for devising effective techniques to determine the optimal number of trees, genetic diversity, and density for producing high-quality, high-yielding seeds. This study, therefore, aims to quantify fertility variation in sandalwood seed production areas and analyze the impact of genetic drift on seed production.
2.1 The study site and observation time
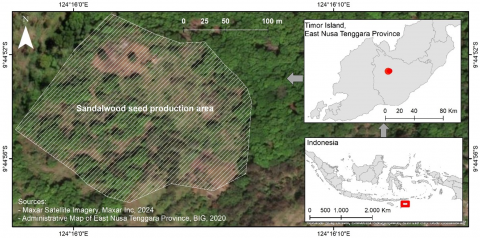
The study was conducted in Netpala Village, Mollo Utara District, Timor Tengah Selatan Regency, East Nusa Tenggara Province, Indonesia, at a 4.09-hectare sandalwood SPA with 259 parent trees. The site’s latitude ranges from 9°44'50" to 9°44'57" S and its longitude ranges from 124°15'57" to 124°15'57" E (Figure 1). Located at 1,056 to 1,090 meters above sea level, the SPA was established in 1992/1993 by the Kupang Forestry Research Institute using genetic material from six provenances on Timor Island: Ajaobaki, Siso, Aenutnanam, Amanuban Barat, Amanatun Selatan, and Kaijob. However, due to missing tree maps and some trees' deaths, the provenance of each parent tree couldn't be determined. As the flowering season of this species occurs twice a year, flower and fruit observations were made during the peak flowering season, which is in December. Flower and fruit production were observed over six months during the peak flowering season, from November 2022 to April 2023.
Figure 1. Study site at the sandalwood seed production area
2.2 Fertility assessment
Fertility is defined as the ability of reproductive traits to produce offspring [33]. Data were obtained by counting the number of flowering and fruiting trees, in addition to the counts of flowers (Flrs) and fruits (Fruits) produced. Using binoculars, cameras, and counters, 24 observations were made at the SPA site every week from November 2022 to April 2023. The fruit set percentage (Fruitset) was calculated using the figures Flrs/Tree and Fruits/Tree for each tree. In addition, the diameter of productive trees was examined to determine its effect on flower and fruit production. To understand fertility dynamics, the data on flowers per tree, fruits per tree, fruitset, and diameter were statistically evaluated.
The fertility of a tree, both male and female, is directly proportional to the quantity of male and female gametes it produces [38]. Fertility was assessed by determining the ratio of reproductive structures (flowers and fruits) produced by an individual tree to the total number produced by all trees. This measurement reflects the contribution of each tree to the overall reproductive output. Gender-specific fertility is determined by estimating the abundance of flowers, representing male reproductive potential, and fruits, representing female reproductive potential, on each tree. Total fertility (pi) is calculated as the average of both male and female reproductive outputs for each tree. Male fertility (mi) and female fertility (fi) represent the i-th tree's relative contributions to reproduction, with mi indicating flower production and fi reflecting seed or fruit output. Total fertility (pi) represents the combined reproductive capacity of the i-th tree in the SPA, encompassing both male and female fertility to measure the tree's overall contribution to population reproduction. If all trees produce the same number of fruits, female fertility is constant at 1/N, where N is the total number of assessed trees, indicating equal contributions to overall reproductive output [22].
2.3 Fertility variation
Fertility variation, defined as differences in the ability of parent trees to produce offspring among individual parent trees, is measured using the sibling coefficient (Ψ). This coefficient quantifies the likelihood that two genes are randomly selected from the same parent's gamete pool compared to equal representation among parents [29]. If Ψ equals 1, each tree contributes equally to the population's gamete gene pool [18, 29, 30]. Ψ is calculated based on the total number of trees evaluated (N) and each tree's individual fertility (pi), which is determined by the estimated number of flowers and/or fruits produced [18, 29, 31].
$\begin{aligned} & \Psi=N \sum_{i=1}^N p_i^2=\sum_{n=1}^N\left(\frac{f_i+m_i}{2}\right)^2= 0.25 N \sum_{n=1}^N\left(f_i^2+m_i^2+2 f_i m_i\right)\end{aligned}$ (1)
In the formula, the variable N represents the total census number of trees being evaluated within the study. The term fi signifies the female fertility of the i-th individual, while mi indicates the male fertility of the same i-th individual. Additionally, pi denotes the probability that two genes in the offspring are derived from the same i-th parent.
2.4 Co-ancestry and population size
Group co-ancestry (θ) assesses the probability that two randomly chosen genes from a population are identical by descent [39], thereby reflecting the level of genetic similarity and relatedness among individuals in that population. When a tree is completely isolated from other trees with no inbreeding, its pair co-ancestry value is zero; conversely, when all trees can self-pollinate, the co-ancestry value reaches 0.5 [40]. To calculate group co-ancestry, the following formula is used [31, 40]:
$\theta=0.5 \sum_{i=1}^N p_i^2=0.5 \sum_{n=1}^N\left(\frac{f_i+m_i}{2}\right)^2$ (2)
Effective population size (Ne) refers to the ideal number of breeding individuals needed to maintain a constant allele frequency across generations in both the parents and their offspring [41]. Ne is a crucial parameter in the study of biological evolution, genetic conservation, and plant breeding, as it helps monitor the rates of genetic drift and inbreeding within populations [42]. In this context, Ne is determined solely by the fertility variation among parents. The Ne value is calculated using the following formula [18]:
$N_e=\frac{N}{\psi}$ (3)
The relative population size (Nr) indicates the ratio of a sampled tree to the overall number of trees within its population [29, 41, 43]. If all parent trees are fertile and have equal fertility (Ψ = 1), Nr equals one, indicating that there is no change in gene frequency between parent trees and their offspring [23].
$N_r=\frac{1}{\Psi}$ (4)
The variance in effective population size (Ne(v)) represents the ideal population size that would produce the same level of sampling variance in allelic frequencies per generation as observed between parent genotypes and their offspring. Ne(v) value is calculated using the following formula [18]:
$N_e^{(v)}=\frac{N}{\Psi-1}$ (5)
Ne(v) has limited relevance for seed production, as it primarily indicates the predicted rate of change in gene frequencies between a parent tree and its offspring, but does not account for all subsequent generations [18].
2.5 Genetic diversity
Genetic diversity (GD) is assessed through group co-ancestry and can be compared between an orchard and a reference population. This study calculates genetic diversity based on fertility, emphasizing the reproductive contributions of individual trees to the overall population's genetic variation. The reference population, from which the orchard trees were selected, is noted for its exceptionally low group co-ancestry [22]. The calculation of genetic diversity uses the formula provided by Varghese et al. [22]:
$G D=1-\theta$ (6)
According to the genetic diversity equation, the co-ancestry group is a component of the genetic diversity that has been depleted since the initiation of tree breeding [18].
3.1 Fertility and variation of fertility
Fertility, which reflects the ability to produce offspring in the next generation, varies significantly based on reproductive traits. The data revealed considerable variation in the number of flowers per tree (Flrs/Tree), fruits per tree (Fruits/Tree), and the fruit set percentage (Fruitset/Tree). Specifically, Flrs/Tree ranged from 60 to 3,575, Fruits/Tree from 2 to 455, and Fruitset/Tree from 0.88% to 13.97%. These variations highlight the diverse reproductive capabilities among trees.
The statistical analysis indicates a significant difference in the number of flowers, fruits, and Fruitset per tree, with a probability value of P < 0.0001. However, no significant correlation was found between tree diameter and the production of flowers, fruits, or Fruitset, with P values greater than 0.05. The average diameter at breast height (dbh) of sandalwood trees in the SPA in ENT is 14.90 cm, ranging from 6.68 cm to 26.74 cm. Variance analysis indicates a significant difference in tree diameter, with a probability value of P < 0.0001.
The results showed that 22.01% of the trees were fertile, with fertility rates ranging from 0.14% to 8.59%. Within the SPA, there were eight trees, constituting 14.04% of the total trees, that exhibited high fertility, each contributing over 5% to flower and fruit production. These eight trees accounted for 50.88% of the total flower production and 60.89% of the total fruit production. Fertility variation within the sandalwood SPA in ENT, as indicated by the sibling coefficient (Ψ), was 2.98.
3.2 Co-ancestry, population size and genetic diversity
Co-ancestry (θ) in sandalwood SPAs was measured at 0.03, reflecting a low level of genetic relatedness among trees. This resulted in an effective population size (Ne) of 19.15, indicating that approximately 19 trees contributed equally to the gene pool, with a genetic diversity (GD) of 0.97.
In sandalwood SPAs in East Nusa Tenggara (ENT), the relative effective population size (Nr) was 0.34, suggesting a limited representation of sampled trees relative to the total population. Additionally, the effective population variance (Ne(v)) was 12.15, indicating a significant potential for genetic drift. This data highlights concerns about genetic stability and the need for management strategies to mitigate genetic drift and maintain genetic diversity.
4.1 Fertility and variation of fertility
Variations in fertility among individual trees in the sandalwood SPAs of East Nusa Tenggara (ENT) are believed to contribute to the low flower production observed, consistent with Kjær's [17] findings that fertility variations lead to reduced flowering. While site conditions also influence flowering habits, Kaosa-ard [44] argued that fertility plays a more significant role. Shorter flowering periods generally result in fewer flowering individuals [45], but the sandalwood SPA in ENT experienced an extended flowering season, with new flowers appearing as late as January. Despite this, only 22.01% of trees produced flowers, a decline from the 38.75% observed in December 2010 [37]. Flower production among the 22.01% of flowering trees varied widely, ranging from 0.14% to 8.59%.
The sandalwood SPA in ENT faces a serious challenge with an exceptionally low fruit set of just 7.18%. The reproductive yield of a tree is significantly affected by the proportion of flowering trees within a stand; as the percentage of flowering trees increases, the overall reproductive output of individual trees also rises, and conversely, a decrease in flowering trees leads to reduced reproductive output [23]. These suboptimal reproductive outcomes are further exacerbated by natural factors, including incompatibility, insufficient cross-pollination, and a scarcity of pollinators, all of which contribute to high rates of flower abortion and low fruit production [12]. Although detailed observations of pollinators were not conducted, bees are likely the primary pollinating insects at the study site. The site's isolation, surrounded by a border of Swietenia macrophylla trees and separated from other sandalwood populations, further discourages pollinators from entering the SPA. This situation severely hinders the SPA's ability to meet the demand for high-quality seeds, especially considering its critical role in supplying genetically superior sandalwood seeds for cultivation across Indonesia. Addressing these reproductive barriers is essential to enhancing the SPA's effectiveness in supporting sustainable sandalwood production.
In the SPA, fertile trees produced an unbalanced number of flowers, limiting the potential for maximum outcrossing between trees. Notably, eight trees, representing just 14.04% of all parent trees, contributed more than 5% each to flower and fruit production. These trees accounted for 50.88% of total flower production and 60.89% of total fruit production, highlighting their significant dominance. Such disparity in gamete contribution from a small number of trees can alter the genetic structure of offspring from this seed source.
The aggregate contribution of trees to overall fertility is represented by the sibling coefficient (Ψ), which reflects the unequal contribution of trees to seed production [23, 29]. If each tree contributes equally to the population's gamete gene pool, then Ψ = 1 [33]. The value of Ψ must be greater than one [34]. In the sandalwood SPAs in ENT, the fertility variation (Ψ) is 2.98, indicating a huge deviation towards inbreeding compared to random mating, where all trees would contribute equally. A Ψ value of 2.98 indicates that the probability of two individuals sharing the same parent is 2.98 times higher compared to a scenario where parental fertility is evenly distributed across the population [34]. Ideally, each tree would contribute 1.75% to total fruit production, but actual contributions range from 0.21% to 3.41%, highlighting substantial fertility variation. Unlike agricultural crop plants, forest trees have large differences in fertility [33]. This high Ψ value is due to both the unequal contribution of fertile trees and the fact that the proportion of infertile trees is notably high at 77.99%, compared to the 22.01% that is fertile trees, further emphasizing the disparity in reproductive success within the SPA. The fertility of trees in sandalwood SPAs is influenced by both genetic factors and the site environment. Environmental factors that exert influence include climate, rainfall, soil type, and seed source management. The 4.09-hectare research site tends to have similar climatic conditions, rainfall, and soil types. Therefore, despite their impact, these elements can be considered consistent throughout the research process. There are currently few silvicultural treatments in sandalwood SPAs, such as soil cultivation, weeding, or fertilization. Intensive silviculture and fertilizer application can increase the number of fertile trees in sandalwood SPAs [46].
Moreover, the occurrence of asynchrony in sandalwood flowering, characterized by maturation of the male and female reproductive organs at different times, frequently results in reproductive failure [16]. Sandalwood flowering asynchrony occurs when male (pollen) and female (ovule) reproductive organs mature at different times [47]. This could result in reproductive failure due to biological mechanisms such as diminished pollination effectiveness. When the stigma is not yet receptive, or when it is already unreceptive by the time pollen is available, the pollination process fails to occur effectively [48-50]. This reduces the chance of male and female gametes interacting. Asynchrony frequently results in the loss of viability of pollen or stigma before they interact with each other. Pollen that is no longer viable cannot produce pollen tubes, while stigmas that are no longer receptive cannot catch pollen [51]. Consequently, the possibility of reproductive failure due to asynchrony may arise in sandalwood [15]. To reduce the deviation towards inbreeding and encourage random mating, the approach is to selectively prune the crowns of very fertile trees, augment the population of fertile trees, and stimulate fertility in less fertile individuals.
4.2 Population size and genetic diversity
The ideal seed source condition is when all trees contribute equally to the gene pool, achieving an effective population size (Ne) of 100%. The sandalwood SPA has a Ne of 19.15 (33.6% of the total number), which suggests that a small fraction of unrelated trees is contributing equally to the gene pool [33]. The number of effective trees is much lower in comparison to other species that utilize intermediate insect pollinators, which exhibit a higher percentage of effective trees, namely 62.5% (Melaleuca cajuputi seedling seed orchard and Tectona grandis clonal seed orchard) [32, 36] and 74.4-78.1% (a seed source established with bulked seeds of superior trees, Azadirachta indica) [30]. However, this number is still comparable to the effective trees in the T. grandis seed production area, which range from 31.2% to 53.5% [23]. This finding emphasizes a notable disparity in reproductive success among the trees. This low Ne is a direct consequence of fertility variation, in which specific trees dominate seed production while others make less or no contribution. Variation in fertility can result in genetic drift, which subsequently decreases the total genetic diversity and heightens the likelihood of inbreeding. Effective management of fertility variation is essential for ensuring the long-term viability of the SPA. The unequal contribution of gametes between trees impacts the genetic diversity of the next generation. A more equitable distribution of contributions from all trees would increase the effective population size, therefore fostering genetic variety and resilience in future generations. This perspective emphasizes the need to implement deliberate measures, such as equally distributed seed harvesting from each tree, to reduce the effects of fertility variation and safeguard the genetic health of the population.
The sandalwood SPA in ENT exhibits worrisome genetic management, characterized by a high fertility variation (Ψ) of 2.98. This variation effect is due to the low relative population size (Nr) of 0.34 and a high effective population size variance (Ne(v)) of 12.15, suggesting a susceptibility to genetic drift. An ideal seed source, with an Nr value of 1, ensures uniform fertility and equal contribution among parent trees, maintaining consistent gene frequency between generations [23]. SPAs often have higher Nr values than natural stands because of targeted management [23]. However, in this study, the presence of several infertile trees, which disturb the gene pool, is likely the cause of the notably lower Nr. This imbalance lowers the value of Ne(v), increasing the likelihood of genetic drift, which has the potential to gradually compromise the SPA's genetic diversity and adaptive ability. To protect the genetic integrity of future generations of sandalwood, management techniques must prioritize improving the fertility and reproductive success of all trees within the SPA. With a co-ancestry score of 0.03, the Sandalwood SPA in East Nusa Tenggara (ENT) maintains a high genetic diversity value of 0.97. In comparison to species utilizing the same insect pollination mechanism (entomophily), this value is somewhat more than the genetic diversity observed in the M. cajuputi seedling seed orchard (0.96) [36] and comparable to the genetic diversity found in the T. grandis clonal seed orchard (0.97) [32]. The high genetic diversity observed can be ascribed to the initial process of selecting parent trees at the founding of the SPA in 1992, which did not take population fertility into consideration. Furthermore, the fertility variation within the SPA is also influenced by the tendency of trees to preserve the phenological patterns of their original provenances [52]. The genetic quality of seeds produced from such seed sources is affected by a combination of factors, including genetic diversity, fertility variation, mating systems, and the combining ability of parent trees [53]. The current study did not consider the gene flow from external populations and the flowering synchronization, which are undoubtedly factors influencing the gene diversity of seeds from the seed stand populations [34].
4.3 Influence of genetic diversity and management considerations for Sandalwood SPA in ENT
Breeding programs are specifically designed to preserve genetic diversity within production populations, a crucial factor for ensuring resilience and adaptability in the face of changing environmental conditions. To effectively achieve this objective, the establishment of conservation stands and clone banks is of paramount importance. For species like sandalwood, which have long life cycles, significant fertility variations, and low fruit production, conserving genetic diversity is particularly important. Effective population size management helps reduce self-pollination and enhances seed production through outcrossing. Therefore, implementing conservation strategies and managing genetic diversity are essential for the long-term success and sustainability of breeding programs [53].
During the domestication process, a decrease in effective population size often leads to reduced genetic diversity, particularly in the first generation. This decline is exacerbated when the contribution of individual parent trees to seed production varies significantly. Ensuring effective outcrossing depends on both the effective contribution of trees and their flowering timing. Sandalwood SPAs in ENT face challenges such as flower abortion and low fruit set, emphasizing the need for more fertile parent trees to enhance seed production. To address this, providing an adequate number of productive trees is a cost-effective approach to improve seed yields and maintain genetic diversity [18, 54].
Sandalwood faces inbreeding depression due to self-pollination, a common adaptation to sexual incompatibility [55]. Low fruit production in sandalwood SPAs is often exacerbated by insufficient pollen, sexual incompatibility, and imbalanced parental contributions. To mitigate the impact of fertility variation, SPA managers should focus on reducing co-ancestry levels by collecting seeds from identified high-fertility trees and mixing them in equal quantities. Collecting an equal number of seeds from each mother tree might artificially balance the population [34]. If increasing genetic diversity through additional trees is not feasible, managing trees with high fertility or pollen production is crucial. It was suggested to mix seeds from various SPAs or different years to mitigate the unequal contribution of gametes among individuals [43, 56]. It is essential to avoid seed collection during periods of low flowering to minimize genetic drift, as high fertility variation during such times can exacerbate genetic drift [17]. Managing these factors effectively is critical for maintaining genetic diversity [22].
Sandalwood SPAs are typically established with a limited number of trees and provenances, focusing primarily on growth rather than fertility. As a result, fruit production from these SPAs tends to be relatively low, largely due to insufficient consideration of the trees' fertility status and opportunities for outcrossing. Incorporating fertility status and diversity into the development of these SPAs is crucial for enhancing plantation productivity. To optimize SPA development and improve fruit production, it would be beneficial to assess and account for the fertility status of the trees, thereby determining the effective number of parents needed at a site.
Genetic diversity is crucial for the successful regeneration of sandalwood plantations. As each generation progresses, a decline in genetic diversity can significantly impact the effectiveness of plantation management. Given that sandalwood has a long life cycle, the effects of selection on genetic diversity become more pronounced over time. Employing low-cost breeding strategies that consider genetic diversity during selection can enhance the quality of future generations. On the other hand, SPA development often prioritizes maximizing growth yield, which can result in variable fertility and inconsistent individual contributions [22]. Focusing solely on growth yield may cause genetic drift, which can negatively impact the quality and adequacy of seeds produced by the seed stand.
Even within the same geographical area, location plays a significant role in the selection of trees, as factors like altitude can influence a tree's ability to flower. Fertility status varies with altitude; for instance, Nikkanen and Ruotsalainen [45] reported that male fertility in Picea abies decreases with increasing altitude. Establishing SPAs across large areas without considering the fertility status of each tree is not ideal, as fertility is inconsistent and highly influenced by both the growing site and the tree's provenance [35]. In Indonesia, it has been observed that even when sandalwood from different provenances is planted together at the same location, each provenance tends to retain the phenological pattern of its original region [52]. Therefore, understanding and accounting for these factors is crucial in selecting suitable trees for SPAs to ensure successful and consistent seed production.
This study’s results and discussion lead to the following conclusions:
1). Sandalwood SPA in ENT has a high fertility.
2). Genetic diversity is generally high as a result of low genetic drift, but the effective population size is small.
3). The result of the study can be used as a scientific basis for the development of seed collection and genetic management strategies.
4). Strategies such as selective pruning of highly fertile tree crowns, increasing the number of fertile trees, and stimulating fertility in less fertile individuals are recommended.
The author would like to thank Martinus Lalus for his assistance in the field data collection, Eko Pujiono for his contribution to creation of the research location map, and all members of the Research Group for Breeding of Wood-Producing Resources at the Research Center for Applied Botany, the Research Organization for Life Science and Environment - National Research and Innovation Agency for their suggestions and input in writing the manuscript. Financial support was provided by the National Research and Innovation Agency and the Faculty of Forestry, Universitas Gadjah Mada.
[1] Brand, J.E., Norris, L.J., Dumbrell, I.C. (2012). Estimated heartwood weights and oil concentrations within 16-year-old Indian sandalwood (Santalum album) trees planted near Kununurra, Western Australia. Australian Forestry, 75(4): 225-232. https://doi.org/10.1080/00049158.2012.10676406
[2] Baldovini, N., Delasalle, C., Joulain, D. (2011). Phytochemistry of the heartwood from fragrant Santalum species: A review. Flavour and Fragrance Journal, 26(1): 7-26. https://doi.org/10.1002/ffj.2025
[3] Jones, C.G., Plummer, J.A., Barbour, E.L. (2007). Non-destructive sampling of Indian sandalwood (Santalum album L.) for oil content and composition. Journal of Essential Oil Research, 19(2): 157-164. https://doi.org/10.1080/10412905.2007.9699250
[4] Pullaiah, T. (2021). Success stories of sandalwood. In: Pullaiah, T., Das, S.C., Bapat, V.A., Swamy, M.K., Reddy, V.D., Murthy, K.S.R. (eds) Sandalwood: Silviculture, Conservation and Applications. Springer, Singapore. https://doi.org/10.1007/978-981-16-0780-6_15
[5] Lingard, K., Perry, M. (2018). An assessment of the regulatory framework of the Western Australian sandalwood industry. Australian Forestry, 81(2): 89-101. https://doi.org/10.1080/00049158.2017.1420455
[6] Burdock, G.A., Carabin, I.G. (2008). Safety assessment of sandalwood oil (Santalum album L.). Food and Chemical Toxicology, 46(2): 421-432. https://doi.org/10.1016/j.fct.2007.09.092
[7] Fatima, T., Srivastava, A., Hanur, V.S., Somashekar, P.V., Rao, M.S. (2019). Genetic diversity estimates of Santalum album L. through microsatellite markers: Implications on conservation. American Journal of Plant Sciences, 10(3): 462-485. https://doi.org/10.4236/ajps.2019.103033
[8] Azeez, S.A., Nelson, R., Prasadbabu, A., Rao, M.S. (2009). Genetic diversity of Santalum album using random amplified polymorphic DNAs. African Journal of Biotechnology, 8(13): 2943-2947. https://doi.org/10.5897/AJB09.381
[9] McLellan, R.C., Dixon, K., Watson, D.M. (2021). Prolific or precarious: A review of the status of Australian sandalwood (Santalum spicatum [R. Br.] A. DC., Santalaceae). The Rangeland Journal, 43(4): 211-222. https://doi.org/10.1071/RJ21017
[10] Arunkumar, A.N., Dhyani, A., Joshi, G. (2019). Santalum album, Indian sandalwood. The IUCN Red List of Threatened Species, 2019: e.T31852A2807668. https://doi.org/10.2305/IUCN.UK.2019-1.RLTS.T31852A2807668.en
[11] Thomson, L.A. (2020). Looking ahead–global sandalwood production and markets in 2040, and implications for Pacific Island producers. Australian Forestry, 83(4): 245-254. https://doi.org/10.1080/00049158.2020.1841441
[12] Ratnaningrum, Y.W., Indrioko, S., Faridah, E., Syahbudin, A. (2017). Gene flow and selection evidence of sandalwood (Santalum album) under various population structures in Gunung Sewu (Java, Indonesia), and its effects on genetic differentiation. Biodiversitas Journal of Biological Diversity, 18(4): 1493-1505. https://doi.org/10.13057/biodiv/d180428
[13] Teixeira da Silva, J.A., Kher, M.M., Soner, D., Page, T., Zhang, X., Nataraj, M., Ma, G. (2016). Sandalwood: basic biology, tissue culture, and genetic transformation. Planta, 243: 847-887. https://doi.org/10.1007/s00425-015-2452-8
[14] Tamla, H.T., Cornelius, J.P., Page, T. (2012). Reproductive biology of three commercially valuable Santalum species: Development of flowers and inflorescences, breeding systems, and interspecific crossability. Euphytica, 184: 323-333. https://doi.org/10.1007/s10681-011-0530-y
[15] Ratnaningrum, Y.W.N., Indrioko, S., Faridah, E., Syahbudin, A. (2018). Population structures and seasons affected flowering, pollination and reproductive outputs of sandalwood in Gunung Sewu, Java, Indonesia. Nusantara Bioscience, 10(1): 12-26. https://doi.org/10.13057/nusbiosci/n100103
[16] Torres, E., Iriondo, J.M., Pérez, C. (2002). Vulnerability and determinants of reproductive success in the narrow endemic Antirrhinum microphyllum (Scrophulariaceae). American Journal of Botany, 89(7): 1171-1179. https://doi.org/10.3732/ajb.89.7.1171
[17] Kjær, E.D. (1996). Estimation of effective population number in a Picea abies (Karst.) seed orchard based on flower assessment. Scandinavian Journal of Forest Research, 11(1-4): 111-121. https://doi.org/10.1080/02827589609382918
[18] Kang, K.S., Lindgren, D. (1998). Fertility variation and its effect on the relatedness of seeds in Pinus densiflora, Pinus thunbergii and Pinus koraiensis clonal seed orchards. Silvae Genetica, 47(4): 196-201.
[19] Muñoz-Gutiérrez, L., Vargas-Hernández, J.J., López-Upton, J., Ramírez-Herrera, C., Jiménez-Casas, M. (2020). Clonal variation in phenological synchronization and cone production in a Pinus patula seed orchard. Silvae Genet, 69: 130-138. https://doi.org/10.2478/sg-2020-0018
[20] Burczyk, J., Chalupka, W. (1997). Flowering and cone production variability and its effect on parental balance in a Scots pine clonal seed orchard. Annales des Sciences Forestières, 54(2): 129-144. https://doi.org/10.1051/forest:19970201
[21] Gömöry, D., Bruchánik, R., Longauer, R. (2003). Fertility variation and flowering asynchrony in Pinus sylvestris: Consequences for the genetic structure of progeny in seed orchards. Forest Ecology and Management, 174(1-3): 117-126. https://doi.org/10.1016/S0378-1127(02)00031-2
[22] Varghese, M., Nicodemus, A., Nagarajan, B., Lindgren, D. (2006). Impact of fertility variation on gene diversity and drift in two clonal seed orchards of teak (Tectona grandis Linn. f.). New Forests, 31: 497-512. https://doi.org/10.1007/s11056-005-2178-8
[23] Varghese, M., Kamalakannan, R., Nicodemus, A., Lindgren, D. (2008). Fertility variation and its impact on seed crops in seed production areas and a natural stand of teak in southern India. Euphytica, 160: 131-141. https://doi.org/10.1007/s10681-007-9591-3
[24] Bila, A.D., Lindgren, D., Mullin, T.J. (1999). Fertility variation and its effect on diversity over generations in a teak plantation (Tectona grandis Lf). Silvae Genetica, 48: 109-114.
[25] Kang, K.S., Bila, A.D., Harju, A.M., Lindgren, D. (2003). Estimation of fertility variation in forest tree populations. Forestry, 76(3): 329-344. https://doi.org/10.1093/forestry/76.3.329
[26] Xie, C.Y., Knowles, P. (1992). Male fertility variation in an open-pollinated plantation of Norway spruce (Picea abies). Canadian Journal of Forest Research, 22(10): 1463-1468. https://doi.org/10.1139/x92-196
[27] Lindgren, D., Prescher, F. (2005). Optimal clone number for seed orchards with tested clones. Silvae Genetica, 54(2): 80. https://doi.org/10.1515/sg-2005-0013
[28] Xie, J., Huang, X., Liu, Y., Zhu, P., Zhu, Y., Li, F., Yao, J., Chen, L., Yang, H. (2022). Variation of fertility and phenological synchronization in Cunninghamia lanceolata seed orchard: Implications for seed production. Forests, 13(10): 1571. https://doi.org/10.3390/f13101571
[29] Kang, K.S., Lindgren, D. (1999). Fertility variation among clones of Korean pine (Pinus koraiensis S. et Z.) and its implications on seed orchard management. Forest Genetics, 6(3): 191-200.
[30] Kamalakannan, R., Varghese, M., Park, J.M., Kwon, S.H., Song, J.H., Kang, K.S. (2015). Fertility variation and its impact on effective population size in seed stands of Tamarindus indica and Azadirachta indica. Silvae Genet, 64: 91-99. https://doi.org/10.1515/sg-2015-0008
[31] Kang, K.S. (2001). Genetic gain and gene diversity of seed orchard crops. Ph.D. Thesis, Swedish University of Agricultural Science, Acta Universitatis Agriculturae Sueciae, Umeå, Sweden
[32] Sumardi, S. (2011). Fertility variation and effective population size in a teak clonal seed orchard. Indonesian Journal of Forestry Research, 8(1): 65-78. https://doi.org/10.20886/ijfr.2011.8.1.65-78
[33] Kang, K.S., Bilir, N., Jeon, K., Kim, Y.J. (2023). Fertility variation and gene diversity in forest populations. Forests, 14(11): 2172. https://doi.org/10.3390/f14112172
[34] Bilir, N., Kang, K.S. (2021). Fertility variation, seed collection and gene diversity in natural stands of Taurus cedar (Cedrus libani). European Journal of Forest Research, 140(1): 199-208. https://doi.org/10.1007/s10342-020-01324-1
[35] Yazici, N., Bilir, N. (2017). Aspectual fertility variation and its effect on gene diversity of seeds in natural stands of Taurus cedar (Cedrus libani A. Rich.). International Journal of Genomics, 2017(1): 2960624. https://doi.org/10.1155/2017/2960624
[36] Kartikawati, N.K. (2016). Fertility variation of Melaleuca cajuputi subsp. cajuputi and its implication in seed orchard management. Indonesian Journal of Forestry Research, 3(2): 83-94. https://doi.org/10.20886/ijfr.2016.3.2.83-94
[37] Sumardi. (2018). Phenology and flowering intensity study of sandalwood (Santalum album Linn.) in Timot Tengah Selatan District - East Nusa Tenggara [Indonesian]. In National Seminar on Biology and Science Education III, 2018, pp. 243-246
[38] Gregorius, H.R. (1989). Characterization and analysis of mating systems. Germany: Ekopan Verlag, Witzenhausen, Germany
[39] Cockerham, C.C. (1967). Group inbreeding and coancestry. Genetics, 56(1): 89-104. https://doi.org/10.1093/genetics/56.1.89
[40] Lindgren, D., Mullin, T.J. (1998). Relatedness and status number in seed orchard crops. Canadian Journal of Forest Research, 28(2): 276-283. https://doi.org/10.1139/x97-217
[41] Kang, K.S., Lindgren, D., Mullin, T.J. (2001). Prediction of genetic gain and gene diversity in seed orchard crops under alternative management strategies. Theoretical and Applied Genetics, 103: 1099-1107. https://doi.org/10.1007/s001220100700
[42] Wang, J., Santiago, E., Caballero, A. (2016). Prediction and estimation of effective population size. Heredity, 117(4): 193-206. https://doi.org/10.1038/hdy.2016.43
[43] Kang, K.S., Kjær, E.D., Lindgren, D. (2003). Balancing gene diversity and nut production in Corylus avellana collections. Scandinavian Journal of Forest Research, 18(2): 118-126. https://doi.org/10.1080/02827580310003696
[44] Kaosa-ard, A. (1996). Teak (Tectona grandis Linn. F.) Domestication and Breeding. UNDP/FAO, Los Banos, Phillipines.
[45] Nikkanen, T., Ruotsalainen, S. (2000). Variation in flowering abundance and its impact on the genetic diversity of the seed crop in a Norway spruce seed orchard. Silva Fennica, 34(3): 205-222. https://doi.org/10.14214/sf.626
[46] Carranca, C., Brunetto, G., Tagliavini, M. (2018). Nitrogen nutrition of fruit trees to reconcile productivity and environmental concerns. Plants, 7(1): 4. https://doi.org/10.3390/plants7010004
[47] Ratnaningrum, Y.W., Indrioko, S. (2015). Response of flowering and seed production of sandalwood (Santalum album linn., Santalaceae) to climate changes. Procedia Environmental Sciences, 28: 665-675. https://doi.org/10.1016/j.proenv.2015.07.078
[48] Raina, M., Kumar, R., Kaul, V. (2017). Stigmatic limitations on reproductive success in a paleotropical tree: Causes and consequences. AoB Plants, 9(4): plx023. https://doi.org/10.1093/aobpla/plx023
[49] Zulkarnain, Z., Eliyanti, E., Swari, E.I. (2019). Pollen viability and stigma receptivity in Swainsona formosa (G. Don) J. Thompson (Fabaceae), an ornamental legume native to Australia. Ornamental Horticulture, 25(2): 158-167. https://doi.org/10.14295/oh.v25i2.2011
[50] Wilcock, C., Neiland, R. (2002). Pollination failure in plants: Why it happens and when it matters. Trends in Plant Science, 7(6): 270-277. https://doi.org/10.1016/S1360-1385(02)02258-6
[51] Tangmitcharoen, S., Owens, J.N. (1997). Pollen viability and pollen-tube growth following controlled pollination and their relation to low fruit production in teak (Tectona grandis Linn. f.). Annals of Botany, 80(4): 401-410. https://doi.org/10.1006/anbo.1996.0440
[52] Ratnaningrum, Y.W., Kurniawan, A. (2019). Floral structure and genetical differences of sandalwood variants in Gunung Sewu (Java, Indonesia), and its effects on breeding systems and reproductive ability. Biodiversitas Journal of Biological Diversity, 20(2): 393-404. https://doi.org/10.13057/biodiv/d200213
[53] Hudson, R., Parent, D. (2016). Forest genetics. Journal of Forestry, 114(2): 84. https://doi.org/10.5849/jof.15-559
[54] Lindgren, D. (2000). Low-intensity tree breeding. In Rapid Generation Turnover into the Breeding Population and Low-Intensity Breeding, 55: 37-48.
[55] Hedegart, T. (1976). Breeding systems, variation and genetic improvement of teak (Tectona grandis L.f.). Tropical Trees, 109-123.
[56] Wojacki, J., Eusemann, P., Ahnert, D., Pakull, B., Liesebach, H. (2019). Genetic diversity in seeds produced in artificial Douglas-fir (Pseudotsuga menziesii) stands of different size. Forest Ecology and Management, 438: 18-24. https://doi.org/10.1016/j.foreco.2019.02.012