Optimization of Somatic Embryo Induction and Nodulation in Mangosteen (Garcinia mangostana L.) Using Carbohydrate Sources and Growth Regulators

Adam Saepudin*![]() | Dedi Natawijaya
| Dedi Natawijaya![]() | Nicky Q. Rachmawati
| Nicky Q. Rachmawati![]() | Tini Sudartini
| Tini Sudartini![]() | Gilang V. Benatar
| Gilang V. Benatar![]()
© 2025 The authors. This article is published by IIETA and is licensed under the CC BY 4.0 license (http://creativecommons.org/licenses/by/4.0/).
OPEN ACCESS
Mangosteen (Garcinia mangostana L.) is a horticultural plant with various beneficial properties from all parts of the plant. However, its cultivation faces several challenges, necessitating the development of efficient propagation techniques. One promising method is the induction of somatic embryos and nodulation. This study aimed to evaluate the effects of honey and sucrose in combination with the growth regulators 2,4-D and BAP on somatic embryo induction and nodulation in mangosteen, determine the optimal treatment concentration, and assess the phases of somatic embryo development and nodular bud formation. The experiment was conducted at the Biotechnology Laboratory, Faculty of Agriculture, Siliwangi University, from September 2022 to February 2023, using a completely randomized design (CRD) with six treatment combinations and four replications. The results showed that all treatments had significant effects (p < 0.05) on the percentage of embryogenic callus, the number of globular-phase somatic embryos, nodules, and nodular buds per explant. The treatment using 5% sucrose + 1 mg L⁻¹ 2,4-D + 3 mg L⁻¹ BAP produced the best results, inducing 29 globular, 4 heart-shaped, and 1 torpedo-stage somatic embryo, along with 30.21 nodules and 3.96 nodular buds per explant. These findings provide a promising protocol for efficient in vitro regeneration of mangosteen, which could support large-scale propagation and conservation efforts.
sucrose, 2,4-D, BAP, mangosteen, nodulation, somatic embryogenesis
Mangosteen (Garcinia mangostana L.) is one of the horticultural crops in the fruit category that is starting to be widely cultivated. In addition to its pleasant taste, the plant—often referred to as the "Queen of Fruits"—is believed to offer various benefits, with industrial applications derived from the fruit, leaves, and rind. Due to its numerous benefits, the demand for mangosteen continues to increase both domestically and internationally. Based on data from the Indonesian Central Bureau of Statistics, in 2020 Indonesia exported 81.15 thousand tons of mangosteen with an export value of US$81.15 million. However, total production was only 322.41 thousand tons—a 30.81% decrease (approximately 75.93 thousand tons) compared to 2019—highlighting the need for improved cultivation practices, especially to increase mangosteen production in order to meet the demand for mangosteen fruit, which is increasing every year [1].
In vitro culture technique is one of the methods used to propagate high-quality mangosteen seeds massively, quickly, and uniformly [2]. Somatic embryogenesis is a process in which somatic cells, including haploid and diploid cells, develop into new plants by undergoing specific embryo development stages without gamete fusion. The study [3] stated that the advantage of this process lies in the genetically uniform propagation results. It can be used for genetic transformation, is virus-free, can regenerate plants from single cells or protoplasts, and can be utilized in synthetic seed technology. Additionally, during the callus induction process, nodular calluses are often formed, which can develop into potential new shoots.
The development of tissue culture systems has been widely explored using various types of explants, media, and plant growth regulators [4]. The success of tissue culture also depends largely on the type of explant and the application of plant growth regulators (PGRs) [5]. According to the study [6], plant growth regulators (PGRs) are organic compounds that play a crucial role in plant growth and development. PGRs regulate the growth rate of individual tissues and integrate these parts to form a structure referred to as a plant [7]. Developmental phases such as globular, heart-shaped, and torpedo stages obtained through somatic embryogenesis resemble the natural embryogenesis process in plants [8]. Various efforts to obtain mangosteen somatic embryos have been made by culturing mangosteen seeds in media containing auxins such as Picloram, 2.4-D, and cytokinins like Thidiazuron [9]. However, the desired formation of somatic embryos has not been achieved. 2.4-D (2,4-Dichlorophenoxy acetic acid) is a potent auxin known to induce callus formation. Its use in the medium is essential to produce embryogenic callus [10]. 2.4-D is commonly used due to its stable nature, making it resistant to damage from light or heat during sterilization [11].
Another factor that can influence somatic embryo formation is the addition of organic materials as a source of carbohydrates, such as sugar, to the culture media. Sucrose is a commonly used sugar type in culture media. Sucrose added to the culture media functions as an energy source because the explants cultured generally have low photosynthetic rates and are not autotrophic [12]. Besides sucrose, honey can be used as another carbohydrate source in culture media. The use of honey in somatic embryogenesis is relatively rare. The study [13] stated that honey, as a carbon source containing carbohydrates, vitamins, and mineral salts, can influence callus growth and cell regeneration. Additionally, the provision of honey as a carbohydrate source can lead to the formation of nodules in mangosteen seed explants. Research [14] on nodule induction in MS media supplemented with honey and BAP either individually or in combination increased the percentage of explants forming nodules and the number of nodules. The addition of 7 mg L-1 BAP resulted in the formation of 28.0 nodules/seed. The purpose of this study is to investigate the effects of using sucrose and honey in combination with plant growth regulators 2,4-D (2,4-dichlorophenoxyacetic acid) and 6-benzylaminopurine (BAP) on somatic embryo induction and nodulation in mangosteen. However, mangosteen propagation remains challenging due to the plant’s long juvenile phase, slow growth, and limited seed viability. Conventional propagation methods are inefficient in meeting the increasing demand for planting material. Therefore, the development of a reliable in vitro propagation protocol, such as somatic embryogenesis, is essential to support large-scale production and conservation efforts.
The study aims to determine the optimal treatment concentrations, identify the phases of somatic embryo induction, and examine the shoots yielded from nodulation results. This study hypothesizes that sucrose and honey, as carbohydrate sources, have different effects on the induction of somatic embryos and nodules in mangosteen when combined with plant growth regulators, with sucrose providing a more defined energy source and honey offering additional bioactive compounds that may enhance cell differentiation and regeneration.
The research was conducted at the Biotechnology Laboratory of the Faculty of Agriculture, Siliwangi University, from September 2022 to February 2023. The plant material used was commercially purchased seeds from ripe fruits of the Puspahiang variety of mangosteen. The mangosteen seeds were separated from the fruit pulp, and uniform-sized seeds were selected for the experiment. The seeds were then sterilized by soaking them in a 20% detergent solution for 10 minutes, followed by a 20% fungicide solution for 30 minutes, and a 30% bayclean solution for 30 minutes. After soaking in each solution, the seeds were rinsed with sterile distilled water three times. Subsequently, the seeds were immersed in 70% alcohol for 10 minutes and rinsed once with sterile distilled water.
Afterward, the planting of the explants was carried out within the Laminar Air Flow Cabinet. The sterilized explants were then cut into 4 pieces horizontally in Petri dishes. The explants were inserted and planted in culture bottles containing pre-prepared growth media. Different treatments involving the addition of carbohydrates and growth regulators were applied using tweezers. The explants were then stored and maintained in an incubation chamber at temperatures ranging from 18℃ to 22℃ with a light intensity of 1000 lux for 24 hours. After subculturing in the 6th week, all explants were kept in the dark until the 14th week, after which the results of the culture were observed every 2 weeks.
This research employed an experimental method structured with a Completely Randomized Design (CRD) consisting of 6 treatments and 4 replications, resulting in 24 experimental units. Each experimental unit comprised 4 culture bottles, with each bottle planted with 4 cross-sectioned pieces of mangosteen seeds. The basal medium used was MS medium (30 g/L sucrose, 4 g L-1 agar, pH 5.8). The treatments included combinations of adding carbohydrate sources and growth regulators, auxin and cytokinin, as follows: A: Honey 3% + 2,4-D 3 mg L-1 + BAP 1 mg L-1; B: Honey 6% + 2,4-D 2 mg L-1 + BAP 2 mg L-1; C: Honey 9 % + 2,4-D 1 mg L-1 + BAP 3 mg L-1 ; D: Sucrose 3% +2,4-D 3 mg L-1 +BAP 1 mg L-1; E: Sucrose 4% +2,4-D 2 mg L-1 +BAP 2 mg L-1; F: Sucrose 5% +2,4-D 1 mg L-1 +BAP 3 mg L-1.
The observations in this study were conducted over a period of 20 weeks, focusing on parameters such as the percentage of callus formation, callus colour and structure, the percentage of embryogenic callus, the number of somatic embryos, the number of nodules formed, and the number of shoots yielded. The observation data were then processed using statistical analysis, entered into the F-test variance table (ANOVA). If significantly different, followed by the DMRT (Duncan Multiple Range Test) test at α = 5%. Histological analysis was also conducted to determine the somatic embryogenesis phases in mangosteen, which cannot be observed directly with the naked eye.
3.1 Callus formation percentage
Callus is an amorphous substance that forms on explants whose cells continuously divide. The appearance of callus on explants serves as an indicator of growth in in vitro culture [15]. The percentage of callus formation is determined to demonstrate the level of responsiveness of explants to the tested treatments. The results of the statistical analysis of callus formation percentage can be seen in Table 1.
The administration of sucrose and honey as carbohydrate sources combined with plant growth regulators (PGRs) 2.4-D and BAP shows significant differences at 28 days after planting (HST culture stage. However, there were no significant differences observed at the 42 HST until 42 days after subculture (HSS) stages regarding the percentage of callus formation. This phenomenon is presumed to occur because the endogenous hormones present in the explants are already sufficient for the mangosteen explant growth. Additionally, the exogenous hormone, 2.4-D auxin, stimulates callus growth in explants faster [16].
However, based on the statistical analysis, treatments D and E significantly influenced the percentage of callus formation at the 28-day after planting stage. This is because the combination of treatments D and E was able to induce callus formation faster and in larger quantities compared to other treatment combinations. The percentage of PGR combination on sucrose application was also higher, ranging from 19.94% to 71.41%, compared to the percentage of callus formation on PGR combination with honey application, which ranged from 13.33% to 59.58%. This finding aligns with the findings of studies [17, 18], indicating that the use of sucrose is more optimal when combined with the auxin 2.4-D. This is because 2.4-D stimulates cell division, while sucrose acts as a provider of carbohydrates for the energy needed in cell division. The application of a high concentration of cytokinin BAP in treatment F did not yield significant differences. This is because the growth-regulating substance that plays a primary role in callus formation is auxin [19, 20] at a higher concentration and equal amounts of cytokinin and auxin.
Energy source for callus growth in the respiration process. These facilities include cell division, enlargement, and the subsequent cell differentiation stage, allowing them to progress effectively.
Table 1. Effect of different carbohydrate sources and plant growth regulators on callus formation percentage at 28 days after planting to 42 days after subculture
|
Treatment |
Callus Formation Percentage (%) |
|
|||
|
28 hst |
42 hst |
14 hss |
28 hss |
42 hss |
|
|
A |
17.92 a |
34.79 a |
44.51a |
55.80 a |
42.81 a |
|
B |
21.61 a |
40.92 a |
46.73 a |
58.05 a |
41.23 a |
|
C |
13.33 a |
42.29 a |
41.98 a |
59.58 a |
46.56 a |
|
D |
28.50 b |
44.08 a |
45.88 a |
52.83 a |
49.52 a |
|
E |
36.04 b |
45.83 a |
53.70 a |
71.41 a |
62.92 a |
|
F |
19.94 a |
39.11 a |
47.02 a |
58.85 a |
45.61 a |
Note: Numbers followed by the same letter are not significantly different based on DMRT test at 95% confidence level; hst: days after planting; hss: days after subculture.
3.2 Color and structure of callus
The color of the callus observed on explants indicates the level of cell division activity visually. The structure of callus is also one of the signs that can be used to determine the quality of callus, indicating whether it is actively dividing or has become stagnant in its cell division process. Callus structure can be categorized into 3 types: friable, non-friable (compact), and intermediate. Good callus has a friable texture, making it easier to separate into individual cells and improving oxygen aeration between cells. In contrast, compact callus structure is difficult to separate due to its dense nature. Intermediate callus, on the other hand, consists of cell clusters, with some being friable and others compact [15]. Visual observations regarding the structure and color of mangosteen callus can be seen in Table 2.
Table 2. Structure and colour of mangosteen callus at culture age 42 days after subculture
|
Treatment |
Callus Structure |
Callus Colors |
|
A |
Friable |
Brownish white |
|
B |
Friable |
Brownish white |
|
C |
Friable |
Brownish yellow |
|
D |
Compact |
Yellowish white |
|
E |
Intermediate |
Yellowish white |
|
F |
Compact |
Greenish yellow |
Based on Table 2, it is evident that treatments A, B, and C exhibit a friable callus structure with a cotton-like white color. Treatment D and F show compact structures with a yellowish- white and greenish-yellow color, respectively, while treatment E demonstrates an intermediate structure with a yellowish- white hue. Friable callus texture is formed due to the presence of endogenous auxin hormones produced internally by the explants forming the callus. Friable callus also undergoes rapid cell division compared to compact callus, as compact callus starts to undergo lignification processes, making its texture harder due to the addition of cytokinin in the form of BAD used for nutrient transport [16]. The formation of compact callus structure occurs during callus induction because of the combined auxin and cytokinin, which influences the water potential within the cells. This leads to increased water absorption from the medium into the cells, resulting in the formation of stiffer cells [21, 22]. Additionally, the treatments D and E, the yellowish-white color of the callus indicates that the cell mass is progressing towards the final stages of active division and is presumed to be embryogenic. Meanwhile, in treatments A and B, the callus appears brownish-white due to the activity of phenolic compounds, causing the callus to turn brownish [15]. This phenomenon leads to a decrease in treatment F, suggesting that chlorophyll formation has begun within the callus cells due to the interaction between 2.4-D and BAP. The change in callus color to greenish white is also a result of light exposure, leading to the formation of chlorophyll in chloroplasts performing photosynthesis [16]. Studies [23-25] also stated that callus turning green indicates an increase in chlorophyll content, white callus that is white to yellowish white is considered embryonic tissue with high starch granule content.
3.3 Embryogenic callus percentage
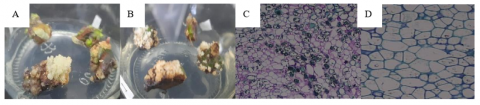
42 days after the planting stage, the previously formed callus started to exhibit signs of embryogenic characteristics. However, these signs did not show a significant effect in each treatment. Embryogenic callus has a compact structure, and the presence of a yellowish-white color indicates embryogenic processes within the callus mass. Additionally, embryonic structures are found within the produced callus [26, 27]. The study [28] also mentions that embryogenic callus has a friable structure, appears yellow, and is solid and nodular, consisting of milk-white embryos. The term “nodular” in somatic embryo induction refers to clear protrusions heading towards the globular phase. Thus, data on suspected embryogenic calluses were collected based on these statements. Subsequently, this was confirmed through histological analysis results, as shown in Figure 1. For instance, in treatment D, the structure of pre-embryo mass (PEM) was observed, characterized by isodiametric shape with large nuclei and dense cytoplasm, which are typical features of cells in embryogenic callus [27]. The statistical analysis results regarding the effect of different carbohydrate sources and plant growth regulators on the percentage of suspected embryogenic callus can be seen in Table 3.
Figure 1. Characteristics of embryogenic and non-embryogenic callus visually and histologically at 42 days after culture: A. Sucrose 3% + 2,4-D 3 mg L-1+ BAP 1 mg L-1 (embryogenic); B. Honey 9% + 2,4-D 1 mg L-1+ BAP 3 mg L-1 (non-embryogenic); C. Sucrose 3% + 2,4-D 3 mg L-1+ BAP 1 mg L-1 (growth and division of embryogenic cells); D. Sucrose 5% + 2,4-D 1 mg L-1 + BAP 3 mg L-1 (non-embryogenic)
Based on the statistical analysis, there is a significant difference between the treatments involving sucrose and honey combined with 2.4-D and BAP at a confidence level of 95% after 14 days of subculture (hss). As shown in Table 3, the application of sucrose with the addition of growth regulators differs significantly at 84 hss compared to honey application in each treatment. This is because sucrose serves as an effective energy source in the culture media, especially for explants that are not autotrophic and have a low photosynthetic rate. Therefore, the carbon source usually obtained from the atmosphere can be fulfilled by sucrose [29, 30]. The addition of honey to the percentage of suspected embryogenic calluses did not significantly impact all combination treatments. This is likely because honey is a complex organic compound containing various other compounds. Explants need to break down these compounds into simpler substances for them to sustain and facilitate plant physiological processes. However, when honey was combined with well-balanced plant growth regulators, such as in treatment B, it resulted in a larger amount of callus compared to other treatments (Treatments A and C), although the yield was not greater than the treatment with added sucrose. Furthermore, at specific concentrations or under saturated conditions, such as in treatment C from 42 days after planting to 98 days after subculture, the addition of higher concentrations of honey as a carbohydrate source could hinder tissue growth and development.
Table 3. Effect of providing different carbohydrate sources and growth regulators on the percentage of embryogenic callus at culture age 42 hst to 98 hss
|
Treatment |
|
|
Percentage of Embryogenic Callus (%) |
|
|
|||
|
42 hst |
14 hss |
28 hss |
42 hss |
56 hss |
70 hss |
84 hss |
98 hss |
|
|
A |
6.25 a |
6.68 a |
7.31 a |
5.56 a |
3.23 a |
2.36 a |
1.44 a |
1.94 a |
|
B |
17.08 a |
4.00 a |
7.07 a |
10.26 a |
8.80 a |
8.23 a |
1.19 a |
2.96 a |
|
C |
2.50 a |
7.34 a |
3.85 a |
2.17 a |
4.23 a |
4.68 a |
2.75 a |
2.36 a |
|
D |
19.17 a |
13.06 a |
15.31 a |
15.33 a |
7.74 a |
7.71 a |
6.26 a |
9.61 a |
|
E |
22.49 a |
20.67 a |
15.61 a |
18.26 a |
10.56 a |
11.26 a |
13.75 b |
9.88 a |
|
F |
14.34 a |
13.86 a |
24.27 b |
35.28 b |
22.63 b |
22.11 b |
15.30 c |
16.27 b |
Note: Numbers followed by the same letter are not significantly different based on DMRT test at 95% confidence level; hst: days after planting; hss: days after subculture.
This aligns with the findings presented by the studies [13, 31], indicating that the addition of relatively high concentrations of sugar as a carbohydrate source in the culture media can inhibit the growth of somatic cells. Sugar, with maintains osmotic pressure, can inadvertently create excessive osmotic pressure, leading to cell death due to cell lysis (rupture of the cell wall). The inadequate formation of embryogenic callus is suspected to be due to the inappropriate addition of auxin and honey concentration. Additionally, in the study [13], honey was added as an organic source rather than a carbohydrate source. Therefore, it is presumed that sucrose was still used as the carbohydrate source in the MS media used in their experiment.
The application of higher sucrose concentration combined with lower auxin concentration, as seen in treatment F, significantly influenced the culture from 28 hss to 98 hss, resulting in the highest percentage of embryogenic callus at 35.28%. This is consistent with the statement made by Lestari [7] that in the formation of embryogenic callus, strong auxins like 2.4-D are commonly used initially. Subsequently, the auxin concentration is reduced, and cytokinins are used in the maturation stage.
3.4 Number of somatic embryos
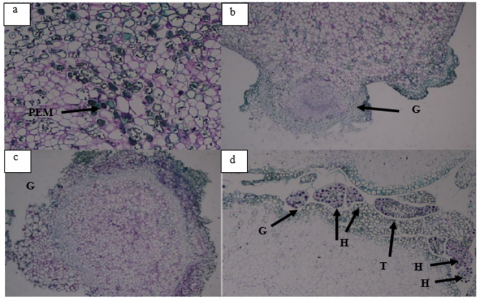
After the formation of embryogenic callus, explants typically start to show the presence of embryos in various stages, including globular, heart, and torpedo stages. The study [20] stated that embryogenic callus has the potential to produce somatic embryos either directly or indirectly, with the latter occurring through callus formation. In this study, histological analysis confirmed the presence of somatic embryos that had reached the heart and torpedo stages (see Figure 2). Visually, the induction of somatic embryos was primarily observed at the globular stage, appearing as round structures with white, cream, and shiny transparent colors (see Figure 3). However, the number of embryos produced was relatively low, as indicated in Table 4. The influence of sucrose and honey combined with 2.4-D and BAP can be observed in Table 5.
Figure 2. Histological stages of mangosteen somatic embryo induction: (a) Early proembryo with 2–3 embryogenic cells (arrow); (b) Globular proembryo with uniform cells; (c) Detached globular stage; (d) Progression to heart and torpedo stages (PEM = pre-embryo mass, G = globular, H = heart, T = torpedo)
Figure 3. Somatic embryos (globular stage) visualized at 84 days after subculture (arrow) in the treatments: (a) Honey 6% + 2,4-D 2 mg L-1+ BAP 2 mg L-1; (b) Sucrose 3% + 2,4-D 3 mg L-1+ BAP 1 mg L-1; (c) Sucrose 5% + 2,4-D 1 mg L-1+ BAP 3 mg L-1
Table 4. The effect of different carbohydrate sources and growth regulators on the number of globular stage embryos at culture ages from 14 hss to 98 hss
|
Treatments |
|
|
Number of Globular Stage Embryos |
|
|
||
|
14 hss |
28 hss |
42 hss |
56 hss |
70 hss |
84 hss |
98 hss |
|
|
A |
0.21 a |
0.38 a |
0.25 a |
0.21 a |
0.59 a |
0.59 a |
0.63 a |
|
B |
0.00 a |
0.19 a |
0.07 a |
0.16 a |
0.64 a |
0.36 a |
0.75 a |
|
C |
0.05 a |
0.05 a |
0.00 a |
0.13 a |
0.59 a |
0.31 a |
0.40 a |
|
D |
0.16 a |
0.23 a |
0.57 c |
0.21 a |
0.84 a |
0.74 a |
1.46 b |
|
E |
0.18 a |
0.19 b |
0.52 b |
0.74 a |
1.38 a |
1.33 b |
1.94 c |
|
F |
0.15 a |
0.41 a |
0.08 a |
1.02 b |
1.43 a |
0.56 a |
1.26 b |
Note: Numbers followed by the same letter are not significantly different based on DMRT test at 95% confidence level; hss: days after subculture.
Table 5. The effect of different carbohydrate sources and growth regulators on the number of nodules at culture ages of 42 hss to 98 hss
|
Treatments |
Number of Nodules |
||||
|
42 hss |
56 hss |
70 hss |
84 hss |
98 hss |
|
|
A |
3.01 a |
3.23 a |
6.67 a |
7.01 a |
7.47 a |
|
B |
5.37 a |
7.09 a |
8.32 a |
8.24 a |
10.44 a |
|
C |
1.60 a |
2.69 a |
4.51 a |
5.58 a |
6.04 a |
|
D |
6.54 b |
6.78 a |
10.29 a |
10.20 b |
11.90 b |
|
E |
12.50 c |
16.19 b |
23.65 b |
24.40 c |
28.15 c |
|
F |
16.50 d |
18.96 b |
25.17 b |
24.83 c |
30.21 c |
Note: Numbers followed by the same letter are not significantly different based on DMRT test at 95% confidence level; hss: days after subculture.
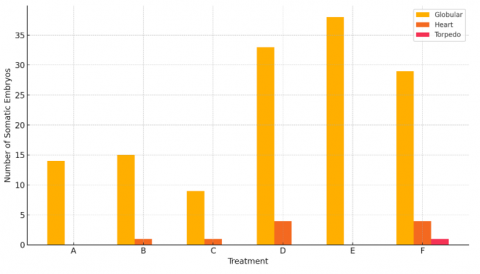
Based on the statistical analysis in Table 4, there is a significant difference in treatments involving the addition of sucrose and honey combined with 2.4-D and BAP at 28 hss, 42 hss, 56 hss, and 98 hss, especially treatments D, E, and F. However, treatments with honey addition did not show significant differences because the number of embryos produced with honey addition was smaller and more uniform compared to sucrose addition. This is consistent with the findings of studies [11, 12], suggesting that explants grow heterotrophically, and without the addition of sucrose, they will not have enough carbohydrates synthesized. The addition of sucrose also provides energy for explant growth, serving as a building material to produce larger molecules necessary for growth. Additionally, autoclaved sucrose is hydrolyzed into glucose and fructose, which can be more efficiently utilized by explants in culture [12]. This demonstrates that different carbohydrate sources can influence the induction process of mangosteen somatic embryos. In contrast, the study [13] resulted in the formation of globular, heart, and torpedo embryonic stages when honey and BAP were used to induce somatic embryos from mangosteen callus from Bengkalis origin. This is likely due to the differences in varieties, the use of different plant growth regulators, and their combinations, all of which can affect the resulting embryo stages.
From 14 days after subculture (hss) to 98 hss, treatment E showed the best results in producing 1.94 globular embryos per explant, totaling 38 globular embryos. However, treatment D, with a higher auxin concentration, was more efficient in producing somatic embryos in the heart stage compared to treatments E and F. According to study [7], this phenomenon occurs because the regeneration through somatic embryogenesis requires several stages involving different concentrations of plant growth regulators. Typically, strong auxins are used in the formation of embryogenic callus, but in subsequent stages, the auxin concentration is reduced, and cytokinins are used during maturation. The addition of 2.4-D is a crucial factor in somatic embryo induction because this auxin, also functioning as a herbicide, can activate the hormones ABA (abscisic acid) and ethylene. Increased ABA hormone levels can induce the expression of proteins that enhance somatic embryogenesis development [32]. Furthermore, the use of different plant growth regulators can influence somatic embryogenesis stages in vitro [12].
In addition to visual observations (Figure 4), histological analysis was conducted to examine tissue development stages experienced by explants, particularly during somatic embryo induction. Histological analysis in tissue culture was performed to understand the developmental phases undergone by the explants, especially during somatic embryo induction. The initial formation of embryos starts with the development of PEM (pre-embryogenic mass) tissue in embryogenic callus. The analysis also aims to identify the forms of each phase achieved, such as globular, heart, and torpedo stages.
Figure 4. Somatic embryo development stages at 98 days of culture
In Figure 2(a), it can be observed that the initial stage of mangosteen somatic embryogenesis involves the emergence of callus cells with distinct nuclei, dense cytoplasm that absorbs strong color, and the presence of starch grains [9]. These cells exhibit embryogenic characteristics with uniformly sized and closely packed cells, isodiametric shapes, one or more large nuclei, thin cell walls, and abundant protoplasm containing relatively small vacuoles [33, 34]. Subsequently, the embryogenic cells swell to form proembryos, further developing into globular embryos as shown in Figure 2(b). In Figure 2(c), the globular stage formed is not connected to the suspensor tissue. The globular protrusion then divides into two parts resembling a heart or scutellar shape, as depicted in Figure 2(d). Additionally, globular, heart, and torpedo stages are simultaneously observed in treatment F.
3.5 Number of nodules per explant
During the induction process of somatic embryos, nodules can form either from callus or emerge directly after subculturing. The influence of different carbohydrate sources and plant growth regulators on the number of nodules can be observed in Table 6, Figure 5, and Figure 6.
Table 6. The effect of different carbohydrate sources and growth regulators on the number of nodular shoots at culture ages of 42 hss to 98 hss
|
Treatment |
Number of Nodular Shoots |
||||
|
42 hss |
56 hss |
70 hss |
84 hss |
98 hss |
|
|
A |
0.00 a |
0.06 a |
0.40 a |
0.24 a |
0.57 a |
|
B |
0.00 a |
0.18 a |
1.31 a |
0.68 a |
0.87 a |
|
C |
0.00 a |
0.13 a |
0.09 a |
0.51 a |
0.50 a |
|
D |
0.32 a |
0.41 a |
0.71 a |
0.98 a |
1.48 a |
|
E |
0.60 a |
1.62 b |
3.88 b |
3.92 b |
4.53 c |
|
F |
0.21 a |
1.13 b |
3.46 b |
2.88 b |
3.96 b |
Note: Numbers followed by the same letter are not significantly different based on DMRT test at 95% confidence level; hss: days after subculture.
Figure 5. Nodule formation at 42 days after subculture in the treatments: (a) Honey 6% + 2,4-D 2 mg L-1 + BAP 2 mg L-1; (b) Sucrose 4% + 2.4-D 2 mg L-1 + BAP 2 mg L-1; (c) Sucrose 5% + 2,4-D 1 mg L-1 + BAP 3 mg L-1
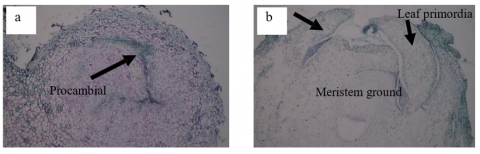
Figure 6. Histology of nodule development at 42 days after subculture in the treatments: (a) Nodule with procambial cells in the meristematic tissue (arrow), sucrose 4% + 2,4-D 2 mg L-1 + BAP 2 mg L-1; (b) Meristematic growth extending from the nodule into the plumule, sucrose 5% +2,4-D 1 mg L-1+ BAP 3 mg L-1
Based on statistical analysis in Table 5, it is evident that the provision of carbohydrates in the form of honey or sucrose combined with 2.4-D and BAP results in significant differences in each treatment from 42 hss to 98 hss, at a 95% confidence level. All treatments involving honey combinations showed no significant differences. However, treatment B displayed higher values in nodule development on explant, reaching 10.44 nodules per explant at 98 hss. This is attributed to the appropriate addition of honey as an organic material, providing water and carbohydrate [35], which facilitates seed growth and development during plant regeneration in processes such as organogenesis, initiated by nodule formation [13], and shoot and leaf formation. Nevertheless, excessively high concentrations of honey as a carbohydrate source can inhibit the growth of new cells [36]. Therefore, treatment B, with a honey concentration of 6% (60 mg L-1 in MS media), was the appropriate composition. If the concentration exceeds 6%, as in treatment C, it could impede the growth of nodules and shoots. Honey also contains high carbohydrates, and when used in excessive amounts in the media, it can hinder nodule growth. This aligns with study [31] that sucrose concentrations commonly used in planting media range from 20 to 60 mg L-1.
In the case of sucrose treatments (D, E, F), there was a significant effect, with treatment F producing the most nodules at 30.21 per explant at 98 days after subculture (hss). This is attributed to the addition of sucrose combined with appropriate plant growth regulators, such as higher concentrations of BAP, which can stimulate the growth of more nodules. According to study [36], the addition of BAP can promote the division and development of meristem cells into nodules on explants, which have the potential to become shoots. The nodules formed can also produce more shoots if further subculturing is performed with suitable media. Figure 6 illustrates the development of the nodules. Unlike globular structures, nodules have elongated procambial cells in the meristem tissue, which then develop into plumules containing the cotyledon and leaf primordia on their right and left sides.
3.6 Number of nodular shoots
During the somatic embryo induction process, shoots can form without going through the embryonic formation process. Shoot formation in mangosteen explants begins with nodule formation, and these nodules swell and develop into shoots around the explant [37]. According to study [38], the nodules formed will develop into shoots, characterized by the emergence of shoot primordia (plumule) and the elongation of cells forming the stem. The transformation of nodules into shoots generally requires the use of different plant growth regulators than in the previous stages. If subcultured into the same medium, the number of nodules and their differentiation into shoots increases. This phenomenon differs from the results observed in treatments involving the combination of honey and sucrose with BAP and 2,4-D. Although the number of shoots fluctuates after 70 days of subculture, subculturing in different concentrations and combinations of media is necessary to obtain more optimal shoot formation. The influence of different carbohydrate sources and plant growth regulators can be seen in Table 6, and the growth of nodules into shoots is illustrated in Figure 7.
Figure 7. Formation of nodular shoots at 84 days after culture in the treatments: (a) Honey 6% + 2.4-D 2 mg L-1 + BAP 2 mg L-1; (b) Sucrose 4% + 2.4-D 2 mg L-1+ BAP 2 mg L-1; (c) Sucrose 5% + 2.4-D 1 mg L-1+ BAP 3 mg L-1
Based on the statistical analysis, there are significant differences in the formation of nodular shoots due to different carbohydrate sources and plant growth regulators. Table 6 shows that honey supplementation did not yield the best results compared to sucrose supplementation in MS media for nodular shoot formation. The study [14] stated that honey supplementation combined with higher concentrations of BAP leads to high osmotic pressure in the medium, disturbing nutrient and exogenous hormone absorption, consequently affecting explant growth. Treatment C, with a higher concentration of BAP (3 mg L-1), produced fewer nodular shoots compared to treatments A (1 mg L-1 BAP) and B (2 mg L-1 BAP). However, treatment B resulted in a higher number of nodular shoots, specifically 1.31 shoots/explant at 70 days after subculture.
This can be attributed to the addition of honey as an organic material at an appropriate concentration, which aids seed growth in plant regeneration, particularly in shoot and leaf formation. The rich mineral composition, vitamins, and other organic compounds present in honey, when added to MS media, are believed to stimulate cell division, leading to shoot formation. The addition of honey in each treatment resulted in fewer shoots compared to the addition of sucrose as a carbohydrate source, combined with different plant growth regulators. This can be observed in treatments D, E, and F, which produced shoots with higher values than treatments A, B, and C. The addition of sucrose can stimulate the growth of more shoots. This aligns with study [37], stating that the addition of sucrose can be utilized as a carbon and energy source, aiding in the growth and development of explant cells since explants are incapable of producing sugars due to not undergoing direct photosynthesis, resulting in low photosynthetic rates and lack of autotrophy in explants cultured in vitro. Additionally, the appropriate addition of plant growth regulators has an impact on the growth of the number of shoots. According to study [13], the concentration of cytokinin added can influence the endogenous hormone content present in the explants. Thus, the proper addition of cytokinin can enhance shoot formation. The use of BAP, an active cytokinin, can promote shoot proliferation, and plants respond well when treated with BAP, making it effective for in vitro shoot production and callus induction [39-41].
The addition of honey in each treatment resulted in fewer shoots compared to the addition of sucrose as a carbohydrate source, combined with different plant growth regulators. This can be observed in treatments D, E, and F, which produced shoots with higher values than treatments A, B, and C. The addition of sucrose can stimulate the growth of more shoots. This aligns with study [36], stating that the addition of sucrose can be utilized as a carbon and energy source, aiding in the growth and development of explant cells since explants are incapable of producing sugars due to not undergoing direct photosynthesis, resulting in low photosynthetic rates and lack of autotrophy in explants cultured in vitro. Additionally, the appropriate addition of plant growth regulators has an impact on the growth of the number of shoots. According to study [13], the concentration of cytokinin added can influence the endogenous hormone content present in the explants. Thus, the proper addition of cytokinin can enhance shoot formation. The use of BAP, an active cytokinin, can promote shoot proliferation, and plants respond well when treated with BAP, making it effective for in vitro shoot production and callus induction [40, 41].
The best treatment was observed in treatment E, which resulted in 4.53 nodular shoots per explant at 70 days after subculture (hss). The application of equal ratios of auxin and cytokinin concentrations yielded good results in nodular shoot growth, although the height growth of the shoots remained relatively slow. This is because shoot growth requires a higher concentration ratio of cytokinin in the cultured media. Moreover, the formation of new nodules continues to occur during nodular shoot development, as indicated in Table 6, showing a continuous increase in the number of nodules from 42 hss to 98 hss. The appropriate concentration of 2,4-D can stimulate cell division and enlargement in plant shoot tips, leading to the growth of new shoots, represented by the nodules. However, excessive auxin concentration can inhibit plant shoot regeneration [41], as seen in treatment D, where the addition of 2,4-D was higher (3 mg L-1) than in treatments E and F, resulting in fewer nodular shoots.
Our findings surpass previous reports, such as study [13], where 9 mL/L honey and 3 mg/L BAP induced only early-stage embryos, whereas our combination of 5% sucrose with 2,4-D and BAP induced more advanced developmental stages (up to torpedo). This indicates that our treatment is more efficient in supporting complete somatic embryogenesis.
The greater efficiency of sucrose may be attributed to its purity and consistency as a disaccharide that provides a stable energy source, while honey—although rich in carbohydrates—contains various enzymes, phenolics, and minerals that may cause oxidative stress or microbial contamination in vitro [23]. Additionally, sucrose promotes more uniform osmotic balance in the medium, which is critical for somatic embryo development.
Excessive BAP levels may inhibit organogenesis by interfering with cell cycle progression, particularly the G1-to-S transition, which is essential for DNA replication and controlled cell division [42]. This supports our observation that lower BAP concentrations, in combination with 2,4-D, facilitated more organized embryogenic development.
Based on the results and discussions, the following conclusions can be drawn:
1. The combination of 5% sucrose with 1 mg/L 2,4-D and 3 mg/L BAP significantly enhanced somatic embryo induction and nodulation in mangosteen explants.
2. Ion process, yielding 29 globular embryos, 4 heart-shaped embryos, and 1 torpedo-shaped embryo. Additionally, it resulted in 30.21 nodules per explant and 3.94 nodular shoots per explant in the mangosteen nodulation process.
3. Future studies should focus on optimizing the maturation phase of somatic embryos and evaluating their conversion into viable plantlets to establish a complete and efficient regeneration system for mangosteen.
Thanks to the Faculty of Agriculture, Siliwangi University, for the Biotechnology Laboratorium facilities that have been provided.
[1] Situmorang, P.S.I., Ayu, S.F., Lindawati. (2023). Market share of Indonesian mangosteen in China market. IOP Conference Series: Earth and Environmental Science, 1241: 012042. https://doi.org/10.1088/1755-1315/1241/1/012042
[2] Engelmann, F. (2010). Use of biotechnologies for the conservation of plant biodiversity. In Vitro Cellular & Developmental Biology - Plant, 47: 5-16. https://doi.org/10.1007/s11627-010-9327-2
[3] Devy, N.F., Hardiyanto, D. (2015). Potential use of in vitro somatic embryogenesis technology for mass propagation of disease-free citrus seeds. Journal of Agricultural Research and Development, 34(4): 169-176.
[4] Ajijah, N., Hartati, R. (2019). Primary and secondary somatic embryogenesis of cacao: The effect of explant types and plant growth regulators. Indonesian Journal of Agricultural Science, 20(2): 69-76. http//doi.org/10.21082/ijas.v.20.n2.2019.p69–76
[5] Agustina, M., Maisura, M., Handayani, R.S. (2020). The effect of different seed cutting treatments and concentrations of BAP for the successful in vitro micrografting of mangosteen (Garcinia mangostana L.). Journal of Tropical Horticulture, 3(1): 1-5.
[6] Gujar, S.B., Pathade, A.G., Pathade, G.R. (2024). The plant growth regulators: Advances, their applications and potential uses in agriculture – A review. Ecology Environment and Conservation, 30(Suppl): 271-280. https://doi.org/10.53550/eec.2024.v30i02s.057
[7] Lestari, E.G. (2011). The role of growth regulators in plant propagation through tissue culture. AgroBiogen Journal, 7(1): 63-66.
[8] Koska, S., Leljak-Levanić, D., Malenica, N., Villi, K., et al. (2025). Developmental phylotranscriptomics in grapevine suggests an ancestral role of somatic embryogenesis. Communications Biology, 8: 265. https://doi.org/10.1038/s42003-025-07712-w
[9] Joni, Y.Z., Prihatini, R., Efendi, D., Roostika, I. (2016). Effect of different sources of plant growth regulator on the induction and development of mangosteen somatic embryos. Indonesian Journal of Agricultural Science, 17(1): 9-16.
[10] George, E.F., Sherrington, P.D. (1984). Plant Propagation by Tissue Culture. Eastern Press, England.
[11] Giszter, R., Fryder, M., Marcinkowska, K., Sznajdrowska, A. (2016). Synthesis, surface properties and biological activity of long chain ammonium herbicidal ionic liquids. Journal of the Brazilian Chemical Society, 27(10): 1774-1781. https://doi.org/10.5935/0103-5053.20160058
[12] Inayah, T. (2015). Effect of sucrose concentration on somatic embryo induction of two peanut (Arachis hypogaea L.) Cultivars in vitro. Jurnal Agribisnis, 9(1): 61-70.
[13] Dreger, M., Mól, R., Deja, A., Raj, E., Mankowska, G., Wielgus, K. (2019). Improved plant regeneration in callus cultures of Sorghum bicolor (L.) Moench. In Vitro Cellular & Developmental Biology - Plant, 55(2): 190-198. https://doi.org/10.1007/s11627-019-09963-9
[14] Reyes-López, D., Leal, E.H., Martínez, C.R.C., Avendaño-Arrazate, C.H., Corona-Torres, T., García-Zavala, J., Ramírez, F.P. (2021). In vitro conservation of vanilla planifolia hybrids in minimal growth conditions. Agro Productividad, 13(12). https://doi.org/10.32854/agrop.v13i12.1773
[15] Rasud, Y. (2020). In vitro callus induction from clove (Syzigium aromaticum L.) leaves on medium containing various auxin concentrations. Indonesian Journal of Agricultural Sciences, 25(1): 67-72.
[16] Inderiati, S., Yanti, F., Mentari, E.R. (2021). Induction of Morphogenic Callus and In Vitro Regeneration of Sugarcane(Saccharum officinarum L. Agriprima Journal of Applied Agricultural Sciences, 5(1): 61-67. https://doi.org/10.25047/agriprima.v5i1.380
[17] Wang, G., Xu, C., Yan, S., Xu, B. (2019). An efficient somatic embryo liquid culture system for potential use in large-scale and synchronic production of Anthurium andraeanum seedlings. Frontiers in Plant Science, 10. https://doi.org/10.3389/fpls.2019.00029
[18] Astuti, R.D., Harahap, F., Edi, S. (2020). Callus induction of mangosteen (Garcinia mangostana L.) in vitro with addition of growth regulators. Journal of Physics: Conference Series, 1485(1): 012029. http//doi.org/10.1088/1742-6596/1485/1/012029
[19] Ge, F., Huang, X., Hu, H., Zhang, Y., et al. (2017). Endogenous small interfering RNAs associated with maize embryonic callus formation. PLOS One, 12(7): e0180567. https://doi.org/10.1371/journal.pone.0180567
[20] Thapa, C.B., Pant, K.K., Bhattarai, H.D., Pant, B. (2022). In vitro induction and proliferation of callus in Piper longum L. through leaf culture. Nepal Journal of Science and Technology, 21(1): 13-22. https://doi.org/10.3126/njst.v21i1.49892
[21] Dantas, L.A., Faria, P.S.A., Dário, B.M.M., Arantes, A.L.M., Silva, F.G., Ávila, R.G., Pereira, P.S., Neto, A.R. (2021). The impact of carbon source on cell growth and the production of bioactive compounds in cell suspensions of Hancornia speciosa Gomes. Scientific Reports, 11: 24315. https://doi.org/10.1038/s41598-021-03845-0
[22] Yıldız, M., Özgen, M. (2004). The effect of a submersion pretreatment on in vitro explant growth and shoot regeneration from hypocotyls of flax (Linum Usitatissimum). Plant Cell Tissue and Organ Culture, 77(1): 111-115. https://doi.org/10.1023/b:ticu.0000016493.03592.c3
[23] Khozin, M. and Mona, M. (2025). Induction and regeneration of tobacco callus using benzyl aminopurine (bap) and furfuryl aminopurine (kinetin) with thin cell layer (tcl) technique. Iop Conference Series Earth and Environmental Science, 1497(1), 012017. https://doi.org/10.1088/1755-1315/1497/1/012017
[24] Wardani D.P., Solichatun, Setyawan, A.D. (2004). Growth and saponin production of Talinum paniculatum Gaertn. callus culture on various addition with 2, 4-dichlorophenoxy acetic acid (2, 4-D) and kinetin. Asian Journal of Natural Product Biochemistry, 2(1): 35-43. https://doi.org/10.13057/biofar/f020106
[25] Shinta, S., Minarno, E.B., Rofiqoh, I. (2020). In vitro embryogenic callus induction of Carica pubescens Lenne and K.Koch using 2,4-D (2,4-dichlorophenoxy acetic acid) and BAP (6-benzylaminopurin). Journal of Biological Researches, 25(2): 38-44. https://doi.org/10.23869/bphjbr.25.2.20205
[26] Al Gabbiesh, A.H., Ghabeish, M., Kleinwächter, M., Selmar, D. (2014). Plant regeneration through somatic embryogenesis from calli derived from leaf bases of Laurus nobilis L.(Lauraceae). Plant Tissue Culture and Biotechnology, 24(2): 213-221. https://doi.org/10.3329/ptcb.v24i2.23554
[27] Indoliya, Y., Tiwari, P., Chauhan, A., Goel, R., Shri, M., Bag, S., Chakrabarty, D. (2016). Decoding regulatory landscape of somatic embryogenesis reveals differential regulatory networks between japonica and indica rice subspecies. Scientific Reports, 6: 23050. https://doi.org/10.1038/srep23050
[28] Hasbullah, N.A., Lassim, M.M., Azis, N.A., Daud, N.F., et al. (2015). Somatic embryo formation in Gerbera jamesonii bolus ex. hook f. in vitro. In International Conference on Agricultural, Ecological and Medical Sciences (AEMS-2015), Phuket, Thailand, pp. 25-28. https://doi.org/10.15242/iicbe.c0415036
[29] Jalil, M., Chee, W.W., Othman, R.Y., Khalid, N. (2008). Morphohistological examination on somatic embryogenesis of Musa acuminata cv. Mas (AA). Scientia Horticulturae, 117(4): 335-340. https://doi.org/10.1016/j.scienta.2008.05.018
[30] Yaseen, M., Ahmad, T., Sablok, G., Standardi, A., Hafiz, I. (2012). Review: Role of carbon sources for in vitro plant growth and development. Molecular Biology Reports, 40(4): 2837-2849. https://doi.org/10.1007/s11033-012-2299-z
[31] Ptak, A., Morańska, E., Skrzypek, E., Warchoł, M., Spina, R., Laurain-Mattar, D., Simlat, M. (2020). Carbohydrates stimulated Amaryllidaceae alkaloids biosynthesis in Leucojum aestivum L. plants cultured in rita®bioreactor. PeerJ, 8: e8688. https://doi.org/10.7717/peerj.8688
[32] Grossmann, K. (2000). Mode of action of auxin herbicides: A new ending to a long, drawn out story. Trends in Plant Science, 5(12): 506-508. https://doi.org/10.1016/S1360-1385(00)01791-X
[33] Nougarède, A. (2003). Analyse d'ouvrage: Meristematic tissues in plant growth and development. Acta Botanica Gallica, 150(2): 239-244. https://doi.org/10.1080/12538078.2003.10515422
[34] Torres, L.F., Diniz, L.E.C., Livramento, K.G.D., Freire, L.L., Paiva, L.V. (2015). Gene expression and morphological characterization of cell suspensions of Coffea arabica L. cv. Catiguá MG2 in different cultivation stages. Acta Physiologiae Plantarum, 37: 175. https://doi.org/10.1007/s11738-015-1924-6
[35] Ketfi, L., Draiaia, R., Necib, A., Mohamadi, N. (2023). Physicochemical and melissopalynological study of some honey samples from the algerian east region. Turkish Journal of Agriculture - Food Science and Technology, 11(4): 780-784. https://doi.org/10.24925/turjaf.v11i4.780-784.5941
[36] Tokuhara, K., Mii, M. (2003). Highly-efficient somatic embryogenesis from cell suspension cultures of phalaenopsis orchids by adjusting carbohydrate sources. In Vitro Cellular & Developmental Biology - Plant, 39(6): 635-639. https://doi.org/10.1079/ivp2003466
[37] Isda, M.N., Amin, N.A., Fatonah, S. (2019). Effect of 6-benzylaminopurine and honey for in vitro shoot initiation of mangosteen seed explant from Riau, Indonesia. Journal of Physics Conference Series, 1351: 012029. https://doi.org/10.1088/1742-6596/1351/1/012029
[38] Xu, L., Cheng, F., Zhong, Y. (2021). Efficient plant regeneration via meristematic nodule culture in Paeonia ostii ‘Feng Dan’. Plant Cell, Tissue and Organ Culture (PCTOC), 149: 599-608. https://doi.org/10.21203/rs.3.rs-986175/v1
[39] Bhau, B.S., Wakhlu, A.K. (2015). A highly efficient in vitro propagation protocol for elephant tusk cactus: Coryphantha elephantidens (Lem.) Lem. Journal of Genetic Engineering and Biotechnology, 13(2): 215-219. https://doi.org/10.1016/j.jgeb.2015.07.003
[40] Swamy, N.R., Ugandhar, T., Praveen, M., Venkataiah, P., Rambabu, M., Upender, M., Subhash, K. (2005). Somatic embryogenesis and plantlet regeneration from cotyledon and leaf explants of Solanum surattense. Indian Journal of Biotechnology, 4(1): 414-418.
[41] Liu, Y., Zhang, H., Guo, H., Xie, L., Zeng, R., Zhang, X., Zhang, S. (2017). Transcriptomic and hormonal analyses reveal that YUC-mediated auxin biogenesis is involved in shoot regeneration from rhizome in cymbidium. Frontiers in Plant Science, 8. https://doi.org/10.3389/fpls.2017.01866
[42] Yang, J., Chen, W., Fan, Y., Zhang, H., Wang, W., Zhang, H. (2016). Ubiquitin protein ligase ring2 is involved in S-phase checkpoint and DNA damage in cells exposed to benzo [a] pyrene. Journal of Biochemical and Molecular Toxicology, 30(10): 481-488. https://doi.org/10.1002/jbt.21811