Antimicrobial and Anti-Biofilm Effects of Essential Oils on Hypermucoviscous Klebsiella pneumoniae

Tabarek G. Ibrahim*![]() | Luma A. Zwain
| Luma A. Zwain![]()
© 2025 The authors. This article is published by IIETA and is licensed under the CC BY 4.0 license (http://creativecommons.org/licenses/by/4.0/).
OPEN ACCESS
The multidrug-resistant bacterium Klebsiella pneumoniae is known to cause critical infections. Its hypermucoviscous phenotype enhances virulence. This study investigates the antimicrobial and anti-biofilm potential of essential oils against these strains. A total of 57 Klebsiella pneumoniae strains were collected from diverse clinical specimens obtained from hospitals in Baghdad. These isolates underwent identification and assessment for hypermucoviscosity characteristics through the string test. The antimicrobial properties of essential oils extracted from thyme (Thymus vulgaris), peppermint (Mentha piperita), and rosemary (Rosmarinus officinalis) were analyzed utilizing both agar diffusion and broth dilution techniques. Biofilm formation and hypermucoviscosity were assessed using the microtiter plate and silicon catheter methods. Among the 57 K. pneumoniae isolates, 9% exhibited the hypermucoviscous phenotype. Resistance was highest to ceftazidime and cefotaxime (94.42%), followed by ampicillin and cefepime (91.42%). Biofilm was formed in 65.46% of the isolates. Essential oils of thyme, peppermint, and rosemary exhibited antimicrobial activity. Notably, thyme and peppermint oils were effective in inhibiting both hypermucoviscosity and biofilm formation. Thyme oil exhibited the highest biofilm inhibition (46.64%), followed by peppermint oil (39.68%). Both oils reduced bacterial adhesion on catheters. The findings of the present study suggest essential oils (Eos), especially those obtained from thyme, peppermint, and rosemary, as potential alternatives for combating hypermucoviscous K. pneumoniae infections, antibiotic resistance, and biofilm-related issues.
hypermucoviscous Klebsiella pneumoniae, essential oil, antimicrobial, anti-biofilm, antiadhesion
Klebsiella pneumoniae is an important intestinal bacterium and opportunistic pathogen implicated in numerous disease conditions. It has gained increasing attention due to its rapid acquisition of antibiotic resistance [1, 2]. Opportunistic infections caused by K. pneumoniae include urinary tract infections (UTIs), pneumonia, sepsis, meningitis, soft tissue infections, diarrhea, and wound infections [3, 4]. Klebsiella pneumoniae is a Gram-negative bacterium that is non-motile and capable of surviving in both oxygen-rich and oxygen-deprived environments. The rising occurrence of multidrug-resistant strains poses a major challenge to global health, as they cause a large number of infections in hospitals with high infection and mortality rates. These bacteria are present in the surface water and soil and on abiotic surfaces such as medical equipment [5]. Klebsiella pneumoniae possesses a variety of virulence factors that enhance its pathogenic ability, such as a polysaccharide capsule (notably serotypes K1 and K2), lipopolysaccharides, iron-scavenging siderophores, fimbrial structures, outer membrane-associated proteins, and the Type VI secretion apparatus (T6SS) [6].
However, highly virulent Klebsiella pneumoniae (HVKP) is more pathogenic than the classical strain (cKp). Since HVKP strains can lead to community-acquired infections, frequently in apparently healthy people, they have become a more serious worldwide hazard than cKp. HVKP may spread quickly in both community and hospital settings because it invades the gastrointestinal system. It is well acknowledged to be a primary cause of pyogenic liver abscesses. These strains are different from cKp in that they can spread to far-off places such as the circulation, lungs, eyes, and nervous system, which can result in infections that are systemic [6-8]. Over the past two decades, a highly virulent form of K. pneumoniae characterized by a hypermucoviscous phenotype has emerged as a significant cause of severe infections [9]. HVKP exhibits a distinctive mucoid phenotype when cultured on agar media, often forming viscous, glossy colonies indicative of increased capsular polysaccharide production. To distinguish it from cKp, HVKP is also referred to as hypermucoviscous (HMV) or hypermucoid K. pneumoniae [8]. Also, most of the major virulence factors of HVKP are usually found on plasmids or mobile genetic elements (MGEs) integrated into the chromosomes, suggesting that horizontal gene transfer (HGT) may be key in the development of the highly virulent phenotype [10, 11]. Since the capsule is a crucial virulence factor, the hypermucoviscous phenotype is usually associated with greater expression of capsule synthesis loci (K-loci) [12].
Essential oils are complex combinations of volatile chemicals, especially terpenes, present extensively in aromatic plants. They readily dissolve in alcohol but remain insoluble in aqueous solutions. EOs are typically a clear and colorless liquid at room temperature, with a distinctive aroma. Their potent taste limits their usage in food, despite their widespread use in the makeup and fragrance industries [13, 14]. Essential oils (EOs), such as those derived from thyme, peppermint, and rosemary, can act as antibiotic adjuvants, enhancing antibiotic activity and serving as synergistic treatments against bacterial infections. They alter the pattern of hypermucoviscosity and have antibacterial activity due to their chemical constituents, especially due to the occurrence of terpenoids [13]. The two most potent terpenoids, thymol and carvacrol, had antibacterial efficacy against a range of infections. The primary constituent of thyme oil, which shares structural similarities with carvacrol, is thymol. The hydroxyl group is located in a different orientation on the phenol ring, whereas thymol also interacts with membrane-bound proteins or the surrounding cytoplasm through hydrophilic and hydrophobic interactions. Carvacrol is a phenolic monoterpenoid compound and a major component of thyme. The antimicrobial activity of carvacrol is similar to that of thymol, which causes functional as well as structural cell membrane damage [15, 16]. In addition to compounds such as menthol, menthone, methyl acetate, limonene, pulegone, menthofuran, β-pinene, eucalyptol, and isomenthone, peppermint essential oil contains other chemicals that contribute as active ingredients against bacterial infections [17].
The ability of essential oils to act against microbes is associated with the mechanisms of action of their constituents within the cell membrane, which leads to membrane dissolution and loss of its integrity due to changes that determine the production of hydrogen, potassium, and calcium ions, resulting in damage to the basic cell survival processes [18]. Rosmarinus officinalis oil possesses distinct hydrophobic properties that disrupt the cytoplasmic in addition to the outer membrane features typical of Gram-negative organisms, as well as the outer layer of plant cells, leading to increased permeability [19, 20]. EOs from Thymus vulgaris, Mentha piperita, and Rosmarinus officinalis play an active role against pathogenic K. pneumoniae strains, influencing the pattern of hypermucoviscosity. These oils are effective in reducing viscosity production and inhibiting biofilm formation in these strains [21]. Carvacrol and thymol reduce biofilm mass by inhibiting the formation of pili, key adhesion structures essential for biofilm development [22]. Thyme oil has a less pronounced effect on the bacterial outer membrane but increases the permeability of the cytoplasmic membrane to ATP, thereby inhibiting biofilm formation [23].
The current investigation sought to evaluate the potential of specific EOs to hamper bacterial growth and the process of formation of biofilm in hypermucoviscous strains of Klebsiella pneumoniae, with a view to identifying potential alternative therapeutic agents for controlling infections caused by this pathogen.
2.1 Bacterial extraction and identification
A collection of fifty-seven (57) identified strains of Klebsiella pneumoniae was obtained from various clinical specimens across hospitals in Baghdad. These included 34 isolates from patients with UTIs, 10 from blood samples, 6 from sputum, and 2 from cerebrospinal fluid. Additional isolates were collected from tissue, artificial bone, burns, and one from a wound. According to the attending specialist physician, all isolates were suspected to be of hospital origin. They were cultured on MacConkey agar, and lactose fermentation was observed. The bacterial isolates were subjected to microscopic examination at 100× magnification. To ascertain their identities, a series of biochemical tests was conducted, including the methyl red, Voges-Proskauer, indole, Simmons citrate, oxidase, and catalase tests, followed by confirmation of the bacterial identification using the VITEK® 2 Compact system.
2.2 Detection of hypermucoviscous bacterial isolates using the string test
Hypermucoviscous bacterial extracts were identified using the string test. A single, circular, and viscous colony grown on MacConkey agar was selected and stretched with a loop. The formation of a sticky filament longer than 5 mm signified a positive result [24].
2.3 Antibiotic resistance test
Antibiotic resistance in Klebsiella pneumoniae isolates was assessed with the VITEK® 2 Compact system equipped with the AST-GN76 card. This system provides rapid and reliable susceptibility results, aligning with Clinical and Laboratory Standards Institute (CLSI) guidelines to ensure accurate and standardized results.
2.4 Biofilm formation assay
Biofilm formation by Klebsiella pneumoniae was assessed using a microtiter plate assay in tryptic soy broth (TSB). A bacterial suspension (1.4 × 10⁸ CFU/mL) was diluted in TSB and added to 96-well plates, with each strain tested in triplicate. Negative controls contained only TSB. Plates were incubated at 37℃ for 48 hours.
After incubation, wells were washed to remove non-adherent cells, then stained with 0.1% crystal violet to visualize biofilms. The excess stain was rinsed off, and the plates were air-dried. Ethanol was used to extract the dye from the biofilms, and the absorbance was measured at 630 nm to quantify biofilm formation [25].
2.5 Extraction of essential oil
Essential oils from Thymus vulgaris (thyme), Mentha piperita (peppermint), and Rosmarinus officinalis (rosemary) were obtained from the College of Science, University of Baghdad. The oils were extracted using a Clevenger-type steam distillation method, where 250 g of dried plant material was boiled in 1.2 L of deionized water for three hours. The extracted oils were collected and stored at 4℃ for later use.
2.6 Evaluation of the antibacterial activity of the EOs against hypermucoviscosity Klebsiella pneumoniae
The antibacterial properties of the essential oils were evaluated against hypermucoviscous K. pneumoniae using the agar well diffusion method. Mueller-Hinton agar plates were coated with a 100 μL bacterial solution (including hypermucoviscous isolates), and wells were made. The wells were then filled with 100 μL of EOs at various doses (50, 25, 12.5, 6.25, and 3.125 mg/ml). Growth inhibition was calculated in millimeters after the plates had undergone the incubation process at 37℃ for 24 hours. Using the serial dilution method, a 100 μL bacterial suspension was added to tubes containing Mueller-Hinton Broth (MHB) supplemented with EOs at the same concentrations. Following incubation at 37℃ for 24 hours, 100 μL of each dilution was put onto plates containing Tryptone Soy Agar, distributed with a glass spreader, and incubated at 37℃ for 24 hours. Next, each dilution's colony count was determined [13].
2.7 Assessment of the effect of EOs on the hypermucoviscosity in K. pneumoniae
The impact of essential oils on hypermucoviscous Klebsiella pneumoniae was evaluated using the string test. Bacterial colonies were cultured on Tryptone Soy Agar (TSA) plates supplemented with thyme, peppermint, or rosemary essential oils at a concentration of 6.5 mg/mL. Control plates lacking essential oils were also prepared. Notably, at 37℃, the entire plates were subjected to incubation for 24 hours. Immediately after that, a sterile loop was used to lift the bacterial colonies, and the length of the resulting mucous thread was measured to assess hypermucoviscosity [13].
2.8 Evaluation of the effect of EOs on biofilm formation by hypermucoviscosity K. pneumoniae
The influence of essential oils on biofilm development by hypermucoviscous K. pneumoniae was investigated employing the silicon Foley catheter model. A sterile 1 cm silicon Foley catheter was autoclaved and then immersed in different concentrations of oils for 3 hours. Following setup, each catheter segment was placed into a 5 mL tube that was sterile with Tryptone Soy Broth (TSB) and inoculated with 100 µL of a hypermucoviscous K. pneumoniae suspension. The bacterial culture was standardized to a turbidity equivalent to 1.5 × 10⁸ CFU/mL using the McFarland method. Tubes were incubated at 37℃ for 48 hours to promote biofilm formation on catheter pieces. After incubation, the catheters were rinsed with phosphate-buffered saline (PBS) to remove loosely attached cells and air-dried at room temperature. Biofilms were stained with 0.1% crystal violet for 15 minutes, then washed with PBS to remove excess dye. To quantify biofilm biomass, 600 µL of 99% ethanol was added to dissolve the retained dye. The ethanol extracts were transferred to a 96-well plate, and absorbance was measured at 630 nm using an ELISA reader [26, 27].
2.9 Effect of EOs on hypermucoviscous K. pneumoniae adhesion to silicone Foley catheter
Essential oils were tested at three concentrations: 50, 25, and 12.5 mg/mL. Segments of silicone Foley catheter (1 cm in length), previously sterilized via autoclaving, were submerged in each concentration of oil, and at 37℃, they were subjected to incubation for 3 hours to allow for absorption. Subsequently, each catheter piece was transferred into sterile tubes containing Tryptone Soy Broth (TSB), followed by inoculation with K. pneumoniae adjusted to a turbidity of 1.5 × 10⁸ CFU/mL using the McFarland standard. Actually, incubation of the setups at 37℃ for 24 hours was to facilitate bacterial adhesion. After incubation, each catheter piece was rinsed in sterile normal saline, and serial dilutions of the wash solution were prepared. Using a sterilized glass spreader, 100 µL of each dilution was plated onto Tryptone Soy Agar (TSA). Afterwards, incubation of the plates at 37℃ for another 24 hours was done, after which bacterial growth was examined and colony-forming units (CFUs) were enumerated.
2.10 Statistical analysis
The Pearson chi-square test was applied to assess the statistical association among the varying proportions of isolates analyzed in the study. A p-value equal to or less than 0.05 was regarded as indicative of statistical significance [28].
3.1 Isolates of K. pneumoniae
Biochemical assays, together with microscopic and culture traits, were used to identify K. pneumoniae isolates. All isolates exhibited growth on MacConkey agar, forming large, circular pink colonies with a shiny, dome-shaped appearance and a mucous texture. All isolates consistently displayed catalase activity and lacked oxidase activity. They were positive in the Simmons citrate and Voges-Proskauer reactions but negative in the indole and methyl red assays. Bacterial identification was confirmed via the VITEK 2 Compact system. The findings indicated statistically significant variations, with p ≤ 0.001 (Table 1).
Table 1. Distribution and proportion of K. pneumoniae isolates by source
|
Percentages |
No. of K. pneumoniae |
No. of Clinical Samples |
Source |
|
58.18a |
32 |
34 |
UTI |
|
18.18b |
10 |
10 |
Blood |
|
10.90c |
6 |
6 |
Sputum |
|
3.63d |
2 |
2 |
Burns |
|
1.81e |
1 |
1 |
Wounds |
|
3.63d |
2 |
2 |
Spinal cord fluid |
|
3.63d |
2 |
2 |
Tissue artificial bone |
|
P-value |
0.001* |
||
* Significant differences at p ≤ 0.001; Similar letters indicate non-significant differences; Different letters indicate significant differences
Table 2. Prevalence and proportion of bacterial isolates exhibiting hypermucoviscosity
|
Number of Isolates |
Hypermucoviscosity > 5 mm |
Hypermucoviscosity < 5 mm |
||
|
Number |
Ratio |
Number |
Ratio |
|
|
55 |
5 |
9% |
50 |
90.90% |
3.2 Prevalence of hypermucoviscous K. pneumoniae
The detection of hypermucoviscous K. pneumoniae revealed that 5 isolates (9%) exhibited the hypermucoviscous phenotype, forming a sticky string greater than five millimeters, considered a positive result in the string test (Table 2).
3.3 Antibiotic resistance profile of K. pneumoniae
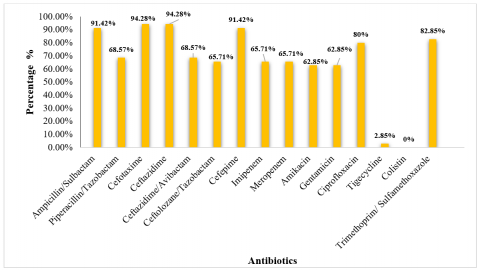
The antibiotic resistance profile of Klebsiella pneumoniae was assessed, and all isolates exhibited resistance to multiple antibiotics. High resistance rates were observed for ampicillin/sulbactam (91.42%), ceftazidime (94.28%), cefotaxime (94.28%), cefepime (91.42%), trimethoprim/sulfamethoxazole (82.85%), and ciprofloxacin (80%). Additionally, resistance to piperacillin/tazobactam, ceftazidime/avibactam, ceftolozane/ tazobactam, imipenem, meropenem, amikacin, and gentamicin ranged from 62.85% to 68.57%. In contrast, low resistance rates were observed for tigecycline (2.85%), while no resistance (0%) was recorded for colistin (Figure 1).
Figure 1. Resistance pattern of K. pneumoniae to antibiotics
Table 3. Biofilm-forming ability of K. pneumoniae
|
Number of Isolates |
Non-formation |
Weak |
Moderate |
Strong |
P-value |
|
55 |
(34.54%)19a |
(63.63%)35b |
(1.81%)1c |
(0%)0c |
0.001* |
* Significant differences at p ≤ 0.001; Similar letters indicate non-significant differences; Different letters indicate significant differences
3.4 Assessment of biofilm development in Klebsiella pneumoniae
Phenotypic detection of biofilm formation using a modified microtiter plate assay revealed that, after 24 hours of incubation in Tryptone Soy Broth (TSB), 34.54% of the isolates showed no biofilm formation, 63.63% exhibited weak biofilm formation, and 1.81% showed moderate biofilm formation. The study demonstrated highly significant differences at a probability level of p ≤ 0.001 (Table 3).
3.5 Antibacterial potential of EO against hypermucoviscous K. pneumoniae
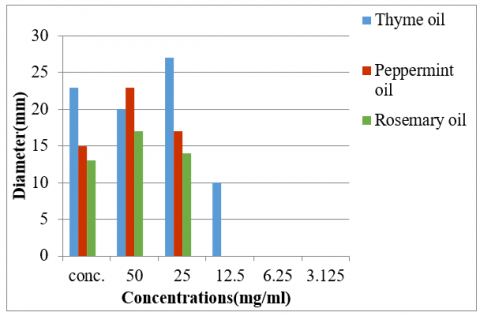
The results obtained from the assay on agar well diffusion, which assessed the effectiveness of EOs against hypermucoviscous Klebsiella pneumoniae, revealed that concentrated thyme, peppermint, and rosemary oils produced 23 mm, 15 mm, and 13 mm, as the respective zones of inhibition. When tested at a concentration of 50 mg/mL, the same oils exhibited inhibition zones of 20 mm, 23 mm, and 17 mm, respectively. At the p< 0.05 level, the study found statistically significant differences (Figures 2, 3, and Table 4). The antibacterial potential of EOs was also evaluated using the serial dilution method. The results showed that the different concentrations (50, 25, 12.5, and 6.25 µL/mL) of EOs inhibited bacterial growth to different extents, as determined by the number of colony-forming units (CFUs). For thyme essential oil, the CFU counts were 110, 240, 220, and 310, respectively. Peppermint essential oil yielded CFU counts of 40, 200, 200, and 250. In contrast, rosemary oil showed higher CFU counts of 270, 380, >3000, and >3000 at the same concentrations, indicating lower antibacterial effectiveness. The control group treated with DMSO exhibited no inhibitory effect on bacterial growth, with CFU counts exceeding 3000. The findings showed that even at minimal levels, thyme and peppermint oils successfully suppressed bacterial growth. In contrast, rosemary oil demonstrated antibacterial potential only at higher concentrations (50 and 25 mg/mL), with no inhibitory effect observed at minimal concentrations (12.5 and 6.25 mg/mL). These findings were statistically significant at the p ≤ 0.05 level (Table 4).

Figure 2. Effect of different concentrations of EOs on hypermucoviscous K. pneumoniae. A: Thymus vulgaris essential oil; B: Mentha piperita essential oil; C: Rosmarinus officinalis essential oil
3.6 Effect of EOs on the hypermucoviscosity in K. pneumoniae
The effect of 6.25 mg/mL of EOs on the inhibition of hypermucoviscosity in Klebsiella pneumoniae was tested using the string test. Thyme oil was the most effective in reducing hypermucoviscosity, as no hypermucoviscosity formation was observed in the string test. Peppermint oil reduced viscosity, with a result of 2 mm compared to the untreated isolates, which showed a viscosity greater than 5 mm. In contrast, rosemary oil had no observable effect on the hypermucoviscosity of the bacteria, as shown in Table 5. The study demonstrated significant differences, with a significance threshold of p ≤ 0.05.
Figure 3. Antibacterial activity of essential oil against hypermucoviscous K. pneumoniae by agar well diffusion method
Table 4. Antibacterial potential of EOs against hypermucoviscous K. pneumoniae using the serial dilution method
|
Essential Oils |
Colony-Forming Unit ×101 |
p-value |
|||
|
mg/ ml |
|||||
|
50 |
25 |
12.5 |
6.25 |
||
|
Thyme oil |
A11a |
B24a |
C22a |
D31a |
0.001* |
|
Peppermint oil |
A4b |
B20b |
B20b |
C25b |
0.01* |
|
Rosemary oil |
A27c |
B38c |
C<300c |
C<300c |
0.05* |
|
Control treatment (DMSO) |
A<300d |
A<300d |
A<300c |
A<300c |
No |
|
P-value |
0.001* |
0.001* |
0.05* |
0.04* |
|
|
0.05 |
|||||
* Significant differences at p ≤ 0.05; Similar lowercase letters in the column indicate non-significant differences; Different lowercase letters in the column indicate significant differences; Similar uppercase letters in a row indicate non-significant differences; Different uppercase letters in a row indicate significant differences.
Table 5. Effect of EOs on hypermucoviscosity in K. pneumoniae
|
Treatment |
String Test Result (mm) |
|
Thyme oil |
0a |
|
Peppermint oil |
2b |
|
Rosemary oil |
<5c |
|
Control treatment (DMSO) |
<5c |
|
Control positive |
<5c |
|
p-value |
0.05* |
* Significant differences at p ≤ 0.001; Similar letters indicate non-significant differences; Different letters indicate significant differences
3.7 Effect of EOs on the biofilm formation of hypermucoviscous K. pneumoniae
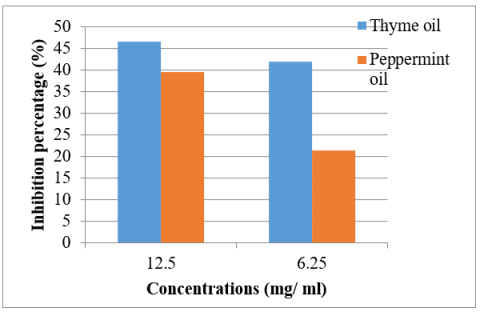
Thyme and peppermint oils were selected to assess their capacity to prevent biofilm development in hypermucoviscous Klebsiella pneumoniae. According to the findings (Figure 4), the percentage of biofilm formation suppression achieved by thyme oil was 46.64% at 12.5 mg/mL and 42.09% at 6.25 mg/mL. Meanwhile, peppermint oil demonstrated an inhibition rate of 39.68% at a concentration of 12.5 mg/mL and 21.52% at a concentration of 6.25 mg/mL. Statistical testing indicated significant differences at the 5% significance level (p < 0.05).
Figure 4. Effect of thyme and peppermint oil on biofilm formation of hypermucoviscous K. pneumoniae using a silicon Foley catheter
3.8 Effect of EOs on adhesion of hypermucoviscous K. pneumoniae to silicon Foley catheter
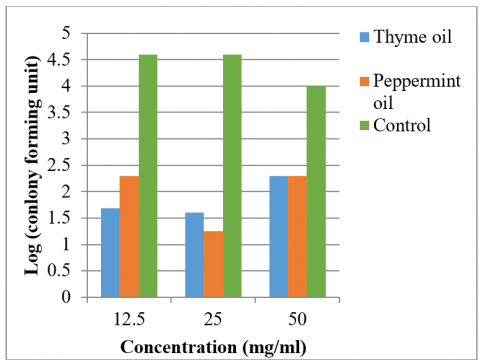
The results demonstrated that thyme and peppermint oils exhibited antiadhesion effects against hypermucoviscous K. pneumoniae on the catheter at concentrations of 50, 25, and 12.5 mg/mL (Figure 5). The number of colonies on the catheter treated with thyme oil were 2×10², 0.04×10³, 0.05×10³ CFU, respectively, while for peppermint oil, the counts were 2×10², 1.8×10¹, 0.2×10³ CFU, respectively. In contrast, the control treatment showed colony counts of 100×10², 40×10³, and 40×10³ CFU, respectively (Table 6). These differences were statistically significant at the p ≤ 0.05 level.
Table 6. Anti-adhesion of essential oils against hypermucoviscous K. pneumoniae
|
Oil |
Colony Forming Unit |
p-value |
||
|
50 mg/mL |
25 mg/mL |
12.5 mg/mL |
||
|
Thyme oil |
A2×102a |
B0.04×103a |
B0.05×103a |
0.05* |
|
Peppermint oil |
A2×102a |
B1.8×101b |
C0.2×103b |
0.001* |
|
Control treatment (DMSO) |
A100×102b |
B40×103c |
B40×103c |
0.04* |
|
p-value |
0.05* |
0.001* |
0.001 |
|
|
0.001* |
||||
* Significant differences at p ≤ 0.05; Similar lowercase letters in the column indicate non-significant differences; Different lowercase letters in the column indicate significant differences; Similar uppercase letters in a row indicate non-significant differences; Different uppercase letters in a row indicate significant differences.
Figure 5. Comparison of the number of colonies adhering to the surface of the silicon Foley catheter before and after treatment
The results indicated that the greatest number of K. pneumoniae isolates were obtained from patients with UTIs, which may be attributed to the higher prevalence of such infections among the patient population. Hasan et al. [29] reported an isolation rate of 35.74% for Klebsiella pneumoniae from UTIs, highlighting it as one of the most dangerous and widespread uropathogens, with high resistance to multiple antibiotics [29]. The current study also observed that the number of isolates from sources other than the urinary tract was lower. This aligns with the findings of Mustafa et al. [30], who reported that bacterial isolates were most prevalent in urine samples (30%), followed by blood (24%), with lower prevalence in sputum and wound samples (18%) and the lowest in burn samples (10%).
Regarding hypermucoviscosity, the results are consistent with the findings of Assefa et al. [31], who reported that 14 out of 39 isolates (35.9%) exhibited hypermucoviscosity. Similarly, Elbrolosy et al. [32] found that 27 out of 84 isolates (32.1%) tested positive in the string test. Liang et al. [33] reported that hypermucoviscosity is linked to the hypervirulence of Klebsiella pneumoniae. Transport and central metabolism genes, as well as mutagenesis mechanisms, impact this phenotype. They further explained that the coexistence of hypermucoviscous and non-hypermucoviscous isolates within the same clonal group may result from mutational mechanisms. Additionally, a disparity in the way metabolism-related genes are regulated and material transport can significantly affect the bacterium’s ability to form biofilms.
Albarri et al. [34] indicated that isolate resistance to antibiotics (piperacillin/tazobactam, cefepime, cefotaxime) was 100% and to antibiotics (ampicillin, meropenem, gentamicin, colistin, cefotaxime, imipenem, trimethyl/sulfa, ciprofloxacin) was 98%, 84%, 72%, 52%, 100%, 34%, 80%, 92%, respectively. Shadkam et al. [35] reported that Klebsiella pneumoniae isolates exhibited resistance to various antibiotics, including trimethoprim-sulfamethoxazole (56.7%), cefotaxime (49.3%), cefepime (52.2%), amikacin (10.4%), imipenem (35.8%), meropenem (41.7%), gentamicin (25.4%), ceftazidime (39.4%), and ciprofloxacin (27.3%). As shown by Mustafa et al. [30], 30% of isolates were ciprofloxacin resistant, 36% of isolates were imipenem resistant, 84% of isolates were ceftazidime resistant, and 80% of isolates were cefepime resistant.
From the present study, the findings indicated that the percentage of bacterial resistance to antibiotics varies over time. This observation is supported by Albarri et al. [34], who reported that antibiotic resistance levels differ across cases, potentially due to variations in the type and number of samples collected, as well as the time and location of sample collection [34]. Shadkam et al. [35] also reported that the geographical distribution of antibiotic resistance varies between countries, with differences attributed to factors such as geographical distance, antimicrobial prescribing patterns in hospitals, and hygiene standards [35]. Additionally, the efflux pump system has been identified as one of the most prevalent antibiotic resistance mechanisms in K. pneumoniae. Studies have shown that these sorts of pumps significantly confer resistance to various antibiotic families, particularly fluoroquinolones [34]. Luna-Pineda et al. [36] further indicated that the hypermucoviscous phenotype of K. pneumoniae strains plays a crucial role in antibiotic tolerance. Proteins, DNA, lipopolysaccharides, and lipids make up the extracellular matrix, which serves as a barrier of protection that prevents antibiotics from penetrating and restricts the creation of novel treatment approaches. Nevertheless, numerous plant extracts have demonstrated promising activity against K. pneumoniae.
In a study conducted by Shadkam et al. [35], the rate of biofilm formation was reported to be 75%, with 31% of isolates forming weak biofilms, 25% forming strong biofilms, and 19% forming moderate biofilms. Similarly, Booq et al. [37] reported that among warm isolates, 39.13% did not form biofilms, 48.83% formed weak biofilms, and 13.04% formed moderate biofilms. Beckman et al. [38] pointed out that most isolates with a high percentage of hypermucoviscosity do not form strong biofilms. During their study, they noted that among the top 15 components of biofilms, only three were mucous, suggesting that these isolates are capable of bypassing the restrictions of increased hypermucoviscosity on biofilm formation. The findings showed that the bacteria's capacity to build biofilms was typically poor, which might be explained by how susceptible they were to growth stimuli.
A recent investigation demonstrated that thyme essential oil possesses strong antibacterial activity against Klebsiella pneumoniae, producing an inhibition zone of approximately 35 mm. This was followed by peppermint oil with a zone of 30 mm and rosemary oil exceeding 25 mm in diameter. The findings highlighted thyme oil as the most potent among the tested essential oils, exhibiting a minimum inhibitory concentration (MIC) of 0.15%. In comparison, peppermint and rosemary oils showed MIC values of 0.60% and 0.45%, respectively [13]. Additionally, research by El-Demerdash et al. [39] reported that thyme oil generated inhibition zones ranging from 14 mm to 32 mm when assessed against K. pneumoniae strains using the agar diffusion technique [39]. Gandova et al. [40] reported that peppermint essential oil exhibited antibacterial activity against K. pneumoniae, producing inhibition zones with a diameter of 2.6 mm. Another study [41] found that the minimum inhibitory concentration (MIC) of rosemary essential oil was determined to be 1.0 mg/mL. The antibacterial properties of essential oils are largely associated with their lipophilic components, particularly bioactive compounds like thymol and carvacrol, which enable them to easily penetrate bacterial cells and disrupt cell membranes, releasing their components into the surrounding environment [42]. Additionally, studies have shown that phenolic compounds like thymol tend to destroy the outer membrane of Gram-negative bacteria. Alturaiki et al. [17] reported that Gram-negative bacteria possess cell walls that are more resistant to chemicals and natural compounds. The fatty polysaccharide layer and the periplasmic space in their cell walls contribute to this resistance, making them less susceptible to antimicrobial treatments compared to Gram-positive bacteria. The study also emphasized that thyme was the most effective treatment against Klebsiella pneumoniae infection. According to their findings, EOs demonstrated significant antimicrobial activity, with inhibition zones greater than 15 mm. The strong antimicrobial effect of EOs is attributed to their complex composition of bioactive compounds and their lipophilic nature. These properties enhance their ability to penetrate biofilms, capsules, and bacterial membranes; disrupt multiple pathways, and aid in the control of various illnesses.
The results of this study demonstrated that hypermucoviscous Klebsiella pneumoniae was affected by EOs. This finding aligns with previous studies on strains with classical viscosity, suggesting that EOs may be effective against both classical and hypermucoviscous forms of K. pneumoniae. Additionally, Dhifi et al. [43] highlighted a key property of EOs, their hydrophobicity. Due to their lipophilic nature, these compounds can integrate into the lipid bilayer of bacterial membranes, altering membrane architecture and increasing permeability. This disruption facilitates the leakage of essential ions and intracellular constituents. Furthermore, the active components in essential oils compromise the integrity of the cytoplasmic membrane, interfering with critical cellular processes such as the proton motive force, electron transport, and active transport systems. Ultimately, these actions lead to the coagulation and collapse of the cell's internal contents. The hypermucoviscous strains of K. pneumoniae were associated with increased antibiotic resistance. The study demonstrated that EOs can reduce hypermucoviscosity across all strains, with some, such as thyme oil, capable of completely eliminating the phenotype. In comparison, rosemary oil showed a lower reduction effect than peppermint oil in mitigating hypermucoviscosity [13]. This observation may imply that bacteria may become more susceptible to environmental influences as a result of the physical barrier produced by the hypermucoviscosity phenotype. EOs appear to disrupt this barrier, weakening the hypermucoviscosity trait and allowing antibiotics to penetrate and act more effectively against the pathogen. Furthermore, combining low concentrations of antibiotics with EOs could provide a viable approach to preventing K. pneumoniae illnesses, even in strains that are multidrug-resistant or exhibit hypermucoviscosity. EOs may alter the hypermucoviscosity phenotype and break through the lipid-soluble barrier that typically hinders antibiotic entry and activity.
Mohamed et al. [44] observed that the acidic water, methanol extract 3 (AWME3), significantly affected hypermucoviscous Klebsiella pneumoniae. All isolates that initially tested positive in the string test showed negative results at an MIC of 0.5 mg/mL. Additionally, the study revealed that phytol, fisetin, and glycitein reduced hypermucoviscosity to the shortest mucosal string length (1 mm) at a concentration of 1 mg/mL. The results showed that a 6.5 mg/mL concentration of thyme and peppermint oils inhibited the formation of hypermucoviscous K. pneumoniae. In contrast, rosemary oil showed no observable effect at the same concentration. This lack of antibacterial activity in rosemary oil may be due to the absence of compounds that target bacterial cells or their virulence factors, particularly those associated with hypermucoviscosity. Reichling [22] reported that thyme essential oil inhibited biofilm formation by 86.1% compared to the control, attributing this strong reduction to the presence of carvacrol and thymol, which interfere with the formation of pili (adhesion molecules). Similarly, Li et al. [45] found that peppermint oil demonstrated a significant ability to inhibit biofilm formation, achieving a 98.2% reduction compared to 95.2% with thyme oil. Mohamed et al. [26] reported that thyme oil exhibited a strong ability to eliminate biofilms, with inhibition rates ranging from 80.1% to 98.0% at a concentration of 10 μL. Their study also confirmed that peppermint oil demonstrated the highest biofilm inhibition among all K. pneumoniae strains, reaching 98.2%, while thyme oil showed a slightly lower inhibition rate of 95.2% [26]. Additionally, Soleimani et al. [23] noted that thyme oil had a moderate effect on the bacterial outer membrane, increasing cytoplasmic membrane permeability to ATP and thereby preventing biofilm formation.
The composition and properties of chemical EOs vary according to plant maturity, biodiversity, geographical region, and growing and treatment conditions affecting their antimicrobial and antibiotic activities [26]. A study reported that type 3 pili contribute to maintaining the structural integrity and fluidity of the polysaccharide capsule in K. pneumoniae [46]. For cells to properly adhere to substrates and for biofilms to develop effectively in this species, fluidity is essential. Type 3 pili promote capsule fluidity by reducing hydrogen bonding between biopolysaccharide molecules. Without them, the capsule forms a denser, more rigid matrix that hinders biofilm formation. The findings showed that, in contrast to prior investigations, the inhibition rate of biofilm growth on the catheter was lower. This may be attributed to the use of hypermucoviscous K. pneumoniae strains in the current study, which have greater antibiotic resistance and antimicrobial agents than classical K. pneumoniae strains.
The results demonstrated a decrease with respect to the number of colonies adhering to the catheter when treated with thyme and peppermint EOs compared to the control group. Mahdi et al. [47] also reported in their study that the adhesion test demonstrated a significant reduction in the adhesion of Gram-negative bacteria when treated with thyme oil, with inhibition rates ranging from 79% to 90%. Džamić et al. [48] reported that thyme oil inhibited the adhesion of E. coli by 39%. Miladi et al. [49] noted that both thyme and rosemary oils exhibited antimicrobial, anti-biofilm, and anti-adhesion activity against the Gram-negative bacterium Salmonella sp., with thyme oil showing a more pronounced effect compared to the other oils. The study proposed that thyme essential oil may act as an effective inhibitor of microbial adhesion to both biotic and abiotic surfaces, ultimately leading to the inhibition of biofilm formation [49]. Khelissa et al. [50] explained that bacterial adhesion to surfaces is influenced by a variety of factors, including surface proteins, pili, and polysaccharides [50].
Flagella contribute significantly to bacterial surface adhesion by overcoming forces that may impede cell-surface communication. In addition to being essential for motility, flagella contribute to surface detection and initiate adhesion, which ultimately leads to biofilm formation. Furthermore, Krukiewicz et al. [51] demonstrated that the inhibitory activity of carvacrol and thymol compounds arises from their ability to reduce pili production, enhance bacterial movement, and downregulate the expression of genes responsible for the production of adhesins and exopolysaccharides (EPS), thus affecting bacterial adhesion. It has been noted that the volatile compounds in both EOs and single oils can influence bacterial membrane integrity. According to the findings, peppermint oil was less effective than thyme oil at preventing bacterial adherence. This could be due to the particular compounds in thyme oil that interfere with key factors required for bacterial adhesion.
EOs from Thymus vulgaris, Mentha piperita, and Rosmarinus officinalis show significant effects on hypermucoviscosity, adhesion, and biofilm formation in hypermucoviscous Klebsiella pneumoniae. Among these, thyme and peppermint oils exhibited the strongest inhibitory activity against these virulence factors, suggesting their potential as alternative antimicrobial agents. For thyme essential oil, the CFU counts were 110, 240, 220, and 310, respectively. Peppermint essential oil yielded CFU counts of 40, 200, 200, and 250.
The authors would like to express their sincere gratitude to all individuals in the Department of Biology, College of Education for Pure Science, Ibn Al-Haitham, University of Baghdad, Iraq, for their invaluable support, expertise, and dedication to this study.
[1] Mohammed, E.A.A.R.K. (2022). Genotype and phenotype investigation of CTX-M gene among multidrug resistant Klebsiella pneumoniae isolates. Iraqi Journal of Biotechnology, 21(2).
[2] Kadum, S.M. (2020). Colistin Susceptibility in Carbapenem Resistant Klebsiella pneumoniae and their ability of biofilm formation. Iraqi Journal of Science, 517-527.
[3] Rahal, B.S., Salman, A.A.A.N.Y., Mohamed, K.K.G.M.M. (2021). The role of EDTA in biofilm eradication of Klebsiella pneumoniae isolated from wound infections. Iraqi Journal of Biotechnology, 1(20).
[4] Al-Salhie, H.H., Al-Kalifawi, E.J. (2020). Antimicrobial and antivirulence activity of magnesium oxide nanoparticles synthesized using Klebsiella pneumonia culture filtrate. Biochemical & Cellular Archives, 20(2).
[5] Khalid, M.Y., Ghareeb, A.M. (2022). Colistin resistant and biofilm formation among multi-drug resistant Klebsiella pneumoniae isolated from different clinical samples. Iraqi Journal of Biotechnology, 21(2): 457-464.
[6] Zhu, J., Wang, T., Chen, L., Du, H. (2021). Virulence factors in hypervirulent Klebsiella pneumoniae. Frontiers in Microbiology, 12: 642484. https://doi.org/10.3389/fmicb.2021.642484
[7] Paczosa, M.K., Mecsas, J. (2016). Klebsiella pneumoniae: Going on the offense with a strong defense. Microbiology and Molecular Biology Reviews, 80(3): 629-661. https://doi.org/10.1128/mmbr.00078-15
[8] Choby, J.E., Howard-Anderson, J., Weiss, D.S. (2020). Hypervirulent Klebsiella pneumoniae–clinical and molecular perspectives. Journal of Internal Medicine, 287(3): 283-300. https://doi.org/10.1111/joim.13007
[9] Prokesch, B.C., TeKippe, M., Kim, J., Raj, P., TeKippe, E.M., Greenberg, D.E. (2016). Primary osteomyelitis caused by hypervirulent Klebsiella pneumoniae. The Lancet Infectious Diseases, 16(9): e190-e195. https://doi.org/10.1016/S1473-3099(16)30021-4
[10] Tian, D., Wang, M., Zhou, Y., Hu, D., Ou, H.Y., Jiang, X. (2021). Genetic diversity and evolution of the virulence plasmids encoding aerobactin and salmochelin in Klebsiella pneumoniae. Virulence, 12(1): 1323-1333. https://doi.org/10.1080/21505594.2021.1924019
[11] Al Ismail, D., Campos-Madueno, E.I., Donà, V., Endimiani, A. (2025). Hypervirulent Klebsiella pneumoniae (hvKp): Overview, epidemiology, and laboratory detection. Pathogens and Immunity, 10(1): 80-119. https://doi.org/10.20411/pai.v10i1.777
[12] Fernández-Manteca, M.G., Ocampo-Sosa, A.A., Vecilla, D.F., Ruiz, M.S., et al. (2024). Identification of hypermucoviscous Klebsiella pneumoniae K1, K2, K54 and K57 capsular serotypes by Raman spectroscopy. Spectrochimica Acta Part A: Molecular and Biomolecular Spectroscopy, 319: 124533. https://doi.org/10.1016/j.saa.2024.124533
[13] Romo-Castillo, M., Flores-Bautista, V.A., Guzmán-Gutiérrez, S.L., Reyes-Chilpa, R., León-Santiago, M., Luna-Pineda, V.M. (2023). Synergy of plant essential oils in antibiotic therapy to combat Klebsiella pneumoniae infections. Pharmaceuticals, 16(6): 839. https://doi.org/10.3390/ph16060839
[14] Abed, I.J., Ahmed, M.E., MH AL-Shimmary, S. (2021). Rosemary volatile oil as a preservative agent in some canned meat foods. Iraqi Journal of Agricultural Sciences, 52(1).
[15] Jafari-Sales, A., Soleimani, H., Moradi, L. (2020). Antibiotic resistance pattern in Klebsiella pneumoniae strains isolated from children with UTIs from Tabriz hospitals. Health Biotechnology Biopharma, 4(1): 38-45.
[16] Al-Wandawy, A.H., Zwain, L.A., Omer, S.A. (2020). Antibacterial and antibiofilm effect of menthol and thymol on vaginal bacteria. Biochemical and Cellular Archives, 20: 3883-3888.
[17] Alturaiki, F.M., Babu Joseph, M.A. (2023). Antibacterial activity of Mentha piperita (Peppermint) oil against wound infections caused by carbapenem-resistant klebsiella in rats. Saudi Journal of Medical and Pharmaceutical Sciences, 9(11): 738-749. https://doi.org/10.36348/sjmps.2023.v09i11.001
[18] Trevisan, S.C.C., Menezes, A.P.P., Barbalho, S.M., Guiguer, É.L. (2017). Properties of Mentha piperita: A brief review. World Journal of Pharmaceutical and Medical Research, 3(1): 309-313.
[19] Alvi, S.S., Ahmad, P., Ishrat, M., Iqbal, D., Khan, M.S. (2019). Secondary metabolites from rosemary (Rosmarinus officinalis L.): Structure, biochemistry and therapeutic implications against neurodegenerative diseases. In Natural Bio-active Compounds, pp. 1-24. https://doi.org/10.1007/978-981-13-7205-6_1
[20] Barreto, H.M., Silva Filho, E.C., Lima, E.D.O., Coutinho, H.D., et al. (2014). Chemical composition and possible use as adjuvant of the antibiotic therapy of the essential oil of Rosmarinus officinalis L. Industrial Crops and Products, 59: 290-294. https://doi.org/10.1016/j.indcrop.2014.05.026
[21] Abdulhasan, G.A. (2015). The biological effect of Rosmarinus officinelis L. essential oil on biofilm formation and some fimbrial genes (fimH-1 and mrkD) of Klebseilla pneumoniae. Iraqi Journal of Science, 56(3C): 2553-2560.
[22] Reichling, J. (2020). Anti-biofilm and virulence factor-reducing activities of essential oils and oil components as a possible option for bacterial infection control. Planta Medica, 86(8): 520-537. https://doi.org/10.1055/a-1147-4671
[23] Soleimani, N., Alkhafaji, A., Mousa, H.M., Ahadi, H. (2023). Antimicrobial activity of thymus vulgaris essential oil against multidrug-resistant Klebsiella pneumoniaee carrying bla NDM gene. Plant, Algae, and Environment, 7(2): 1127-1138. https://doi.org/10.48308/jpr.2024.234105.1058
[24] Hyun, M., Lee, J.Y., Kim, H.A. (2024). Clinical and microbiologic analysis of Klebsiella pneumoniae infection: Hypermucoviscosity, virulence factor, genotype, and antimicrobial susceptibility. Diagnostics, 14(8): 792. https://doi.org/10.3390/diagnostics14080792
[25] Lordelo, R., Branco, R., Gama, F., Morais, P.V. (2024). Assessment of antimicrobial resistance, biofilm formation, and surface modification potential in hospital strains of Pseudomonas aeruginosa and Klebsiella pneumoniae. Heliyon, 10(9): e30464. https://doi.org/10.1016/j.heliyon.2024.e30464
[26] Mohamed, S.H., Mohamed, M.S., Khalil, M.S., Azmy, M., Mabrouk, M.I. (2018). Combination of essential oil and ciprofloxacin to inhibit/eradicate biofilms in multidrug‐resistant Klebsiella pneumoniae. Journal of Applied Microbiology, 125(1): 84-95. https://doi.org/10.1111/jam.13755
[27] Namasivayam, S.K.R., Christo, B.B., Arasu, S.K., Kumar, K.A.M., Deepak, K. (2013). Anti biofilm effect of biogenic silver nanoparticles coated medical devices against biofilm of clinical isolate of Staphylococcus aureus. Global Journal of Medical Research, 13(3): 1-7.
[28] SAS. (2004). Statistical analysis system, user’s guide. Statistical version 9.1. SAS Institute.
[29] Hasan, T.H., Alasedi, K.K., Aljanaby, A.A.J. (2021). A comparative study of prevalence antimicrobials resistance Klebsiella pneumoniae among different pathogenic bacteria isolated from patients with urinary tract infection in Al-Najaf City, Iraq. Latin American Journal of Pharmacy, 40: 174-178.
[30] Mustafa, M.S., Abdullah, R.M. (2020). Role of oqxA and oqxB genes in the development of multidrug resistant phenotype among clinical Klebsiella pneumoniae isolates from various cases. Iraqi Journal of Science, 61(8): 1902-1912.
[31] Assefa, M., Amare, A., Belachew, T., Tigabu, A. (2024). Hypermucoviscous Klebsiella Pneumoniae caused community-acquired pneumonia in Gondar, northwest Ethiopia. Clinical Laboratory, 70(9): 1630-1636.
[32] Elbrolosy, A., Eissa, N., Al-Rajhy, N., El-Mahdy, E., Mostafa, R. (2021). Characterization of virulence genetic profile and resistance patterns of clinical Klebsiella pneumoniae isolates: Classic versus hypermucoviscous phenotypes. Microbes and Infectious Diseases, 2(3): 516-528.
[33] Liang, Q., Chen, N., Wang, W., Zhang, B., et al. (2024). Co-occurrence of ST412 Klebsiella pneumoniae isolates with hypermucoviscous and non-mucoviscous phenotypes in a short-term hospitalized patient. Msystems, 9(7): e00262-24. https://doi.org/10.1128/msystems.00262-24
[34] Albarri, O., AlMatar, M., Var, I., Köksal, F. (2024). Antimicrobial resistance of clinical Klebsiella pneumoniae isolates: Involvement of AcrAB and OqxAB efflux pumps. Current Molecular Pharmacology, 17(1): E310323215266. https://doi.org/10.2174/1874467217666230331081434
[35] Shadkam, S., Goli, H.R., Mirzaei, B., Gholami, M., Ahanjan, M. (2021). Correlation between antimicrobial resistance and biofilm formation capability among Klebsiella pneumoniae strains isolated from hospitalized patients in Iran. Annals of Clinical Microbiology and Antimicrobials, 20: 1-7. https://doi.org/10.1186/s12941-021-00418-x
[36] Luna-Pineda, V.M., Rodríguez-Martínez, G., Salazar-García, M., Romo-Castillo, M. (2024). Plant-origin components: new players to combat antibiotic resistance in Klebsiella pneumoniae. International Journal of Molecular Sciences, 25(4): 2134. https://doi.org/10.3390/ijms25042134
[37] Booq, R.Y., Abutarboush, M.H., Alolayan, M.A., Huraysi, A.A., et al. (2022). Identification and characterization of plasmids and genes from carbapenemase-producing Klebsiella pneumoniae in Makkah Province, Saudi Arabia. Antibiotics, 11(11): 1627. https://doi.org/10.3390/antibiotics11111627
[38] Beckman IV, R.L., Cella, E., Azarian, T., Rendueles, O., Fleeman, R.M. (2024). Diverse polysaccharide production and biofilm formation abilities of clinical Klebsiella pneumoniae. npj Biofilms and Microbiomes, 10(1): 151.
[39] El-Demerdash, A.S., Alfaraj, R., Farid, F.A., Yassin, M.H., Saleh, A.M., Dawwam, G.E. (2024). Essential oils as capsule disruptors: Enhancing antibiotic efficacy against multidrug-resistant Klebsiella pneumoniae. Frontiers in Microbiology, 15: 1467460. https://doi.org/10.3389/fmicb.2024.1467460
[40] Gandova, V., Fidan, H., Iliev, I., Lasheva, V., Stankov, S., Stoyanova, A., Yavorov, N. (2023). Comparative analysis of antibacterial activity and physicochemical properties of peppermint and cornmint essential oils and their main compound menthol. Journal of Chemical Technology and Metallurgy, 58(4): 664-671.
[41] Hassan, A.A., Iskander, D., Oraby, N.H. (2022). Evaluation of the synergistic antimicrobial activities of selenium nanoparticles and rosemary oil against Aspergillus fumigatu and Klebsiella pneumoniae recovered from respiratory infection in cattle in Giza Governorate, Egypt. Exploratory Animal & Medical Research, 12(1): 24-32. https://doi.org/10.52635/eamr/12.1.24-32
[42] Taher, M.S., Salloom, Y.F., Al-Asadi, R.A.U.H., Al-Mousswi, Z.J., Alamrani, H.A. (2021). The medicinal importance of Thyme plant (Thymus vulgaris). Biomedicine, 41(3): 531-534.
[43] Dhifi, W., Bellili, S., Jazi, S., Bahloul, N., Mnif, W. (2016). Essential oils’ chemical characterization and investigation of some biological activities: A critical review. Medicines, 3(4): 25. https://doi.org/10.3390/medicines3040025
[44] Mohamed, H., Marusich, E., Divashuk, M., Leonov, S. (2024). A unique combination of natural fatty acids from Hermetia illucens fly larvae fat effectively combats virulence factors and biofilms of MDR hypervirulent mucoviscus Klebsiella pneumoniae strains by increasing Lewis acid–base/van der Waals interactions in bacterial wall membranes. Frontiers in Cellular and Infection Microbiology, 14: 1408179. https://doi.org/10.3389/fcimb.2024.1408179
[45] Li, L., Gao, X., Li, M., Liu, Y., et al. (2024). Relationship between biofilm formation and antibiotic resistance of Klebsiella pneumoniae and updates on antibiofilm therapeutic strategies. Frontiers in Cellular and Infection Microbiology, 14: 1324895. https://doi.org/10.3389/fcimb.2024.1324895
[46] Wang, H., Wilksch, J.J., Strugnell, R.A., Gee, M.L. (2015). Role of capsular polysaccharides in biofilm formation: an AFM nanomechanics study. ACS Applied Materials & Interfaces, 7(23): 13007-13013. https://doi.org/10.1021/acsami.5b03041
[47] Mahdi, F., Barka, M.S., Bendahou, M., Ouslimani, S., Benziane, M.Y., Muselli, A. (2024). In vitro inhibitory effect of four essential oils against fluoroquinolone-resistant enterobacteriaceae responsible for community-acquired urinary tract infections. Tropical Journal of Natural Product Research, 8(8): 8233.
[48] Džamić, A.M., Nikolić, B.J., Giweli, A.A., Mitić‐Ćulafić, D.S., Soković, M.D., Ristić, M.S., Knežević‐Vukčević, J.B., Marin, P.D. (2015). Libyan Thymus capitatus essential oil: Antioxidant, antimicrobial, cytotoxic and colon pathogen adhesion‐inhibition properties. Journal of Applied Microbiology, 119(2): 389-399. https://doi.org/10.1111/jam.12864
[49] Miladi, H., Mili, D., Slama, R.B., Zouari, S., Ammar, E., Bakhrouf, A. (2016). Antibiofilm formation and anti-adhesive property of three mediterranean essential oils against a foodborne pathogen Salmonella strain. Microbial Pathogenesis, 93: 22-31. https://doi.org/10.1016/j.micpath.2016.01.017
[50] Khelissa, S.O., Abdallah, M., Jama, C., Faille, C., Chihib, N.E. (2017). Bacterial contamination and biofilm formation on abiotic surfaces and strategies to overcome their persistence. Journal of Materials and Environmental Sciences, 8(9): 3326-3346.
[51] Krukiewicz, K., Kazek-Kęsik, A., Brzychczy-Włoch, M., Łos, M.J., Ateba, C.N., Mehrbod, P., Ghavami, S., Shyntum, D.Y. (2022). Recent advances in the control of clinically important biofilms. International Journal of Molecular Sciences, 23(17): 9526. https://doi.org/10.3390/ijms23179526