Maria Kanan*![]() | Herawati
| Herawati![]() | Inda Widiarti Hafid
| Inda Widiarti Hafid![]() | Sandy Novryanto Sakati
| Sandy Novryanto Sakati![]() | Mihwan Sataral
| Mihwan Sataral![]() | Muhammad Syahrir
| Muhammad Syahrir![]()
© 2025 The authors. This article is published by IIETA and is licensed under the CC BY 4.0 license (http://creativecommons.org/licenses/by/4.0/).
OPEN ACCESS
Although flies are conventionally regarded as vectors of infectious agents, increasing evidence suggests that their associated microbiota may harbour bioactive secondary metabolites with therapeutic potential. In this study, the antimicrobial properties of microbial isolates obtained from fly specimens were investigated, with a focus on three strains: Bifidobacterium minimum (A2), Bacillus sp. (A3), and Brucella melitensis biovar (A4). Secondary metabolites were extracted and partitioned into ethyl acetate-soluble and insoluble fractions, followed by chemical characterisation using thin-layer chromatography (TLC) to determine relative fluorescence (RF) values and preliminary compound classification. Bioactivity screening was conducted via the disk diffusion assay on Mueller–Hinton Agar (MHA) against a panel of clinically relevant, drug-resistant pathogens, including Escherichia coli, Staphylococcus aureus, Klebsiella pneumoniae, Pseudomonas aeruginosa, Acinetobacter baumannii, Salmonella typhi, and Candida albicans. Phytochemical analysis revealed the presence of flavonoids, terpenoids, phenolics, and alkaloids, compounds known for their antimicrobial properties. Differential inhibition profiles were observed: B. minimum exhibited modest activity against S. aureus, Bacillus sp. demonstrated inhibitory effects on both E. coli and K. pneumoniae, while B. melitensis biovar displayed the broadest spectrum of activity, with a maximum inhibition zone of 14 mm recorded against A. baumannii. The presence of bioactive secondary metabolites in fly-derived microbial communities underscores their underexplored potential as sources of novel antimicrobial agents. These findings contribute to the growing interest in exploiting symbiotic or commensal microbiota from unconventional ecological niches for the discovery of compounds active against multidrug-resistant pathogens, addressing a pressing need in contemporary antimicrobial drug development.
antimicrobial metabolites, fly-associated microbiota, secondary metabolites, drug-resistant pathogens, microbial inhibition, disk diffusion assay
Microorganisms play a crucial role in maintaining ecological balance, both as pathogenic agents and as normal microflora that support various organisms [1, 2]. As pathogens, microorganisms can cause diseases in plants, animals, and humans [3, 4]. Meanwhile, their role as normal microflora is vital in digestive processes, vitamin synthesis, and protection against infections, thereby contributing to homeostasis [5]. A comprehensive understanding of microorganisms is essential for designing effective health management strategies [6, 7]. In addition, various microbial sources, including those found in insects such as flies, offer significant opportunities in the discovery of potential bioactive compounds [8, 9]. These microorganisms possess a unique ability to produce a wide range of chemical compounds that can act as antibiotics, anticancer agents, or antimicrobial substances [10, 11]. Previous studies have isolated seven bacterial strains capable of producing antimicrobial compounds from Musca domestica and Chrysomya megacephala [12]. The unique potential of the fly microbiome in the development of novel antimicrobial agents lies in its highly complex and dynamic microbial community, which has adapted to harsh and competitive environments, thereby increasing the likelihood of producing rare or previously unidentified bioactive metabolites [9, 13].
However, the specific characteristics and effectiveness of these active compounds against antibiotic-resistant bacteria remain largely unknown. Research on microorganisms isolated from fly bodies holds great promise for pharmaceutical and biotechnological developments [8, 9]. Moreover, the diversity of microbial communities within insect ecosystems provides a unique environment for discovering bioactive compounds that may be absent in conventional sources [14]. Recent studies have increasingly emphasized the antimicrobial properties of fly-associated microbiota, which highlight the isolation of novel antimicrobial peptides and secondary metabolites from fly gut symbionts. In addition, research revealed the presence of phenolic compounds with strong antibacterial activity produced by endosymbiotic bacteria from Lucilia sericata. Similarly, a study identified a new group of alkaloid compounds from microorganisms isolated from the surface of Calliphora vicina, demonstrating potential as inhibitors of multidrug-resistant bacteria. These findings reinforce the hypothesis that flies are not only vectors of disease but also promising biological reservoirs in the search for novel antimicrobial compounds [15, 16].
Bioactive compounds produced by microorganisms exhibit significant antimicrobial activity, indicating their potential for the development of new therapeutic agents [17-19]. This activity is derived from various classes of compounds, including peptides, alkaloids, and fatty acids, each with distinct mechanisms of inhibiting pathogenic microorganisms [20]. The application of these bioactive compounds in healthcare is especially relevant in light of the growing global challenge of antibiotic resistance [21]. Several fly species are recognized as vectors of various pathogens, making them valuable subjects in microbiological and public health research [7, 22, 23]. Their role in transmitting bacteria, viruses, and parasites contributes to the spread of infectious diseases among humans and animals [24-26]. Understanding the role of flies in pathogen ecosystems can inform the development of more effective and sustainable strategies to prevent disease transmission [27-29].
The potential of microorganisms found on flies as sources of active compounds remains largely underexplored [21, 30]. This knowledge gap may be attributed to the challenges in isolating and characterizing the complex microbial communities inhabiting fly bodies, as well as limited understanding of their ecological interactions [14, 21]. Therefore, a focused exploration of the fly microbiome may reveal novel antimicrobial agents that are overlooked in conventional environments, especially given the increasing urgency to address multidrug-resistant pathogens [31]. An interdisciplinary approach that integrates microbiology, biochemistry, and analytical technologies is essential to thoroughly investigate this potential [32, 33]. This study aims to isolate and characterize the active compounds produced by these microorganisms and to evaluate their antibacterial activity against both pathogenic and antibiotic-resistant bacteria. The findings are expected to contribute to the development of new antimicrobial agents to combat antibiotic resistance and to highlight the promise of microbe-derived compounds as viable alternatives to conventional antibiotics.
2.1 Processing of the sample and bacterial isolates
This study utilized bacterial isolates from flies known for their potential to produce antimicrobial compounds, as identified in prior research. The sample comprised three bacterial isolates: Bifidobacterium minimum [NR 044692.2] (A2), Bacillus sp. [MH071158.1] (A3), and Brucella melitensis bv [NR_076080.2] (A4). Additionally, pathogenic bacterial isolates were included as targets for antimicrobial testing, specifically clinically significant microorganisms such as Escherichia coli, Staphylococcus aureus MRSA, Klebsiella pneumoniae, Pseudomonas aeruginosa, Acinetobacter baumannii, Salmonella typhi, and Candida albicans.
2.2 Rejuvenation of bacterial isolates
To prepare Nutrient Agar (NA) medium, dissolve the NA powder in distilled water and heat the solution to boiling. Sterilize the medium in an autoclave at 121℃ for 15 minutes, then pour it into sterile Petri dishes and allow it to solidify. Bacterial isolates known for producing antimicrobial compounds, specifically B. minimum, Bacillus sp., and B. melitensis bv, were inoculated onto the surface of the Nutrient Agar using sterilized inoculation loops under aseptic conditions. Following inoculation, the Petri dishes were incubated at 37℃ for 24 hours, a temperature conducive to the growth of various bacteria and the development of bacterial colonies. Subsequently, the isolates were fermented to facilitate the production of antimicrobial compounds.
2.3 Production of antimicrobial compounds
The production of antimicrobial compounds begins with the preparation of the Nutrient Broth (NB) media. This involves dissolving NB powder in distilled water according to the specified composition on the packaging, followed by sterilization in an autoclave at 121℃ for 15 minutes. Once sterilization is complete, the media is cooled to room temperature to ensure it is ready for use. The fermenter is then sterilized to maintain aseptic conditions. After all preparations are finalized, bacterial isolates are inoculated into the sterile Nutrient Broth. This medium is incubated at 37℃ for 24 hours. Following incubation, the inoculum is transferred to the fermenter at a ratio of 5-10% of the total media volume. This inoculation is a critical step in the production process, as it initiates the bacteria's production of antimicrobial compounds once introduced into the fermenter.
2.4 Fermentation process
The fermentation process commenced with the establishment of parameters in the fermenter, setting the temperature to 37℃, agitation to 100-200 rpm, and continuous aeration at a rate of 1-2 L/min for aerobic bacteria. These conditions are designed to optimize bacterial growth and activity throughout the seven-day fermentation period. After this duration, the fermenter was deactivated, and aeration ceased. At this point, bacterial biomass was separated from the fermentation medium via centrifugation, with the resulting supernatant containing the antimicrobial compounds produced during fermentation.
2.5 Extraction of active compounds
The extraction of active compounds begins with separating the fermented bacterial culture from the media through centrifugation. The resulting supernatant containing the active compounds is then subjected to further extraction. This supernatant is mixed with ethyl acetate in a separatory funnel at a ratio of 1:1 to 1:2 and shaken for 5-10 minutes to ensure thorough mixing. The ethyl acetate phase is subsequently separated, and this extraction process is repeated 2-3 times to enhance the recovery of active compounds. Once the ethyl acetate phase is collected, it is transferred to a round flask for evaporation using a rotary evaporator, which removes the solvent and leaves a concentrated extract of active compounds. This thick extract is stored in sterile vials at low temperatures (4℃ or -20℃) to prevent degradation. The next step involves purifying the active compounds.
2.6 Purification of active compounds
The fermentation products from isolates A2, A3, and A4 were placed into separate separatory funnels, into which ethyl acetate and water were added at a 1:1 ratio. This ratio was adjusted depending on the volume of the fermentation mixture. The mixture was gently stirred and allowed to stand until two distinct layers formed, with the ethyl acetate phase rising to the top due to its lower density than that of water. The tap on the separatory funnel was then opened to separate the two phases, which were collected into containers. The ethyl acetate-soluble (upper phase) and ethyl acetate-insoluble (lower phase) extracts from A2, A3, and A4 were evaporated in a water bath at 60℃ to obtain concentrated extracts. The concentrated ethyl acetate-soluble extracts from A2, A3, and A4 were dissolved in a chloroform:methanol (1:1, v/v) mixture, while the ethyl acetate-insoluble extracts were dissolved in methanol. A small volume of each ethyl acetate-soluble and -insoluble extract was applied to a TLC plate using a capillary tube. The TLC chamber was prepared with an eluent of n-hexane and ethyl acetate in a 9:1 ratio. The TLC plate was then inserted vertically into the chamber, which was sealed tightly to allow the eluent to ascend the silica gel plate. After removal from the chamber, the plate was allowed to dry at room temperature and then examined under UV light at 254 and 366 nm to visualize the spots.
2.7 Antibacterial activity test
Antibacterial effectiveness was assessed using the agar diffusion method in sterile Petri dishes. Test bacterial isolates, including E. coli, S. aureus, K. pneumoniae, P. aeruginosa, A. baumannii, S. typhi, and C. albicans, were cultured on MHA using the swab technique. Sterile paper disks were then immersed in the compound fractions and placed on the surface of the solidified MHA. The plates were incubated at 37℃ for 24 hours. Ethyl acetate was used as the negative control. After incubation, the antibacterial activity was determined by measuring the diameter of the inhibition zones around the disks using a digital caliper or millimeter ruler. The absence or presence and size of these clear zones indicated the level of antimicrobial effectiveness of the tested fractions compared to the negative control. Each experiment was performed in triplicate (n = 3), and data analysis was conducted without statistical testing.
3.1 Visualization and characterization of active compounds
Observation of the TLC plate under a UV lamp at 254 nm revealed dark patches of compounds against a green background (Figure 1(A)). This indicates that the detected compounds can absorb UV light at this wavelength. These findings provide crucial preliminary insights into the presence and characteristics of the compounds in the sample, serving as an initial step for further analysis of the active compounds produced.
Observation of the TLC plate under a UV lamp at 366 nm revealed that the separated compound patches appeared luminous or fluorescent, providing further insights into their properties (Figure 1(B)). This fluorescence confirms the compounds' presence and offers clues regarding their chemical characteristics and molecular structure. Spraying the TLC plate with FeCl3 solution elicited a notable reaction with phenolic compounds, resulting in blue or purple coloration in the detected spots, indicating their presence (Figure 1(C)). The application of Citroborate solution produced a specific color reaction that aids in identifying flavonoid compounds in the sample (Figure 1(D)). Dragendorff solution was used to detect alkaloids, with the resulting spots exhibiting yellow to orange coloration, clearly indicating their presence (Figure 1(E)). Finally, spraying Vanillin sulfate solution on the TLC plate imparted a yellow-brown color to the separated compounds, signifying the presence of terpenoids in the sample (Figure 1(F)).
Characterization of active compounds in microbial isolates from flies revealed the presence of phenolics, flavonoids, alkaloids, and terpenoids (Figure 2). The RF values were consistent across the three isolates, with notably high signals for flavonoids and phenolic compounds. Isolate A4 exhibited stronger fluorescence for terpenoids and alkaloids (RF = 0.8), indicating a richer metabolite profile than the others.
Figure 1. Visualization of active compounds produced by fly-associated microbial isolates
The TLC schematic illustrates the migration behavior of four major metabolite classes. Compounds with higher polarity, such as flavonoids and phenolics, showed the furthest migration (RF = 0.9), whereas terpenoids and alkaloids migrated slightly less (RF = 0.8). These results are in agreement with the fluorescence-based characterization and validate the diversity of bioactive constituents.
Figure 2. RF values of active compounds in each microbial isolate associated with flies
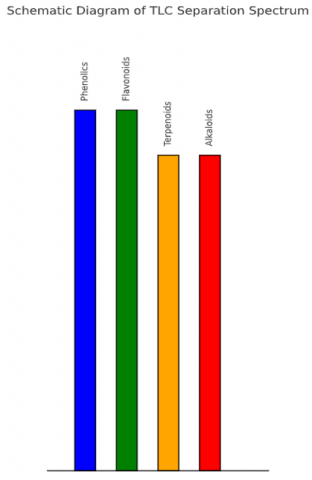
Following the RF profile shown in Figure 2, a schematic representation of the TLC separation spectrum is illustrated in Figure 3.
3.2 Antibacterial activity test for each isolate
The antibacterial activities of ethyl acetate-soluble extracts from B. minimum (A2), Bacillus sp. (A3), and B. melitensis (A4) were evaluated using the disk diffusion method on agar media against seven clinically relevant pathogenic microorganisms. The results showed varying inhibition zones among the isolates, indicating differential susceptibility of the pathogens to the respective bioactive compounds (Figure 4 and Table 1).
Figure 3. Schematic diagram of the TLC separation spectrum of bioactive compounds produced by fly-associated microbial isolates
Figure 4. Antibacterial activity (inhibition zone diameters) of ethyl acetate-soluble extracts from B. minimum (A2), Bacillus sp. (A3), and Brucella melitensis (A4) against seven clinically relevant pathogenic microorganisms
Table 1. Inhibition zone activity of B. minimum (A2), Bacillus sp. (A3), and B. melitensis (A4) isolates on seven types of pathogenic microorganisms
|
Pathogenic Microorganisms |
Average Zone of Inhibition (mm) |
|||||
|
Ethyl Acetate Soluble Extract |
Ethyl Acetate Insoluble Extract |
|||||
|
A2 |
A3 |
A4 |
A2 |
A3 |
A4 |
|
|
E. coli |
0 |
7 |
11.5 |
0 |
0 |
0 |
|
P. aeruginosa |
0 |
0 |
12.5 |
0 |
0 |
0 |
|
C. albicans |
0 |
0 |
13 |
0 |
0 |
0 |
|
S. aureus MRSA |
8 |
7 |
10 |
0 |
0 |
0 |
|
A. baumanii |
0 |
0 |
14 |
0 |
0 |
0 |
|
S. typhi |
0 |
0 |
7 |
0 |
0 |
0 |
|
K. pneumonia |
0 |
12 |
7 |
0 |
0 |
0 |
Figure 5. Disk diffusion assay images showing inhibition zones formed by ethyl acetate-soluble extracts of B. minimum, Bacillus sp., and B. melitensis against seven pathogenic microorganisms
Among the three isolates, B. melitensis (A4) demonstrated the most comprehensive antibacterial profile, exhibiting inhibitory activity against all tested pathogens. The largest inhibition zones were recorded against Acinetobacter baumannii (14 mm) and Candida albicans (13 mm). Bacillus sp. (A3) showed moderate activity, particularly against Escherichia coli (7 mm), Staphylococcus aureus MRSA (7 mm), and Klebsiella pneumoniae (12 mm). In contrast, B. minimum (A2) displayed limited antibacterial activity, with measurable inhibition only against Staphylococcus aureus MRSA (8 mm), suggesting a narrow spectrum of activity. Importantly, none of the ethyl acetate-insoluble fractions exhibited any detectable antibacterial activity, indicating that the bioactive compounds responsible for inhibition were present exclusively in the ethyl acetate-soluble phase.
Figure 4 presents a quantitative bar graph illustrating the average inhibition zone diameters, enabling direct comparison of antibacterial efficacy across isolates and pathogens, whereas Figure 5 displays photographic images of the agar plates, visually confirming the presence, shape, and clarity of the inhibition zones formed by each extract.
Together, these figures provide complementary perspectives: while Figure 4 facilitates analytical comparison based on quantitative data, Figure 5 offers qualitative visual validation, thereby enhancing the overall interpretation of the antibacterial potential demonstrated by each isolate.
Characterization of active compounds in fly-associated microorganisms has identified a diverse range of secondary metabolites. The isolates B. minimum (A2), Bacillus sp. (A3), and B. melitensis (A4) exhibited distinct metabolite profiles, reflecting the unique compounds produced by each species. Isolates A3 and A4 produced flavonoids, terpenoids, phenolics, and alkaloids, known for their antioxidant and antimicrobial properties [34-36], while isolate A2 did not yield any alkaloids. All three isolates demonstrated high RF values for flavonoids and phenolic compounds, suggesting significant potential for therapeutic applications. These compounds have been previously recognized for their role in inhibiting the growth of pathogenic bacteria [37-39]. Mechanistically, flavonoids are known to disrupt microbial membrane integrity and inhibit nucleic acid synthesis, whereas alkaloids often interfere with protein function and bacterial cell wall biosynthesis [40, 41].
Isolate A4 demonstrated the capacity to produce secondary metabolites, specifically terpenoids and alkaloids, with high RF values. These findings suggest a diverse polarity in the metabolites produced by this isolate, offering potential flexibility in targeting antibiotic-resistant pathogens [42]. Previous studies have indicated that terpenoids and alkaloids from bacterial isolates possess significant antibacterial properties [41, 43]. For instance, diterpenoids isolated from Streptomyces spp. and alkaloids from Actinomycetes have shown potent activity against MRSA and ESBL-producing bacteria in other reports, highlighting the therapeutic potential of similar compound classes [44, 45].
As resistance to conventional antibiotics continues to rise, the exploration of these compounds as therapeutic alternatives is increasingly pertinent. The antibacterial activity test results indicated that the insoluble fraction of the ethyl acetate extract did not produce inhibition zones against the tested pathogenic microorganisms. This suggests that active compounds with antimicrobial potential may either be absent in this fraction or not sufficiently dissolved to interact with the target microorganisms. Another possibility is that the compounds are bound within a polymeric or hydrophobic matrix that inhibits their diffusion, or their bioavailability is reduced due to low solubility in aqueous environments [46, 47]. The active compounds might also be trapped within a complex matrix that prevents their release by the solvents, thereby diminishing their effectiveness. Further investigation using fractionation techniques and bioassay-guided isolation is warranted to explore whether these inactive fractions contain cryptic or latent bioactive compounds [48, 49].
The antibacterial activity of B. minimum (A2), Bacillus sp. (A3), and B. melitensis (A4) isolates against seven pathogenic microorganisms revealed varying zones of inhibition for the ethyl acetate soluble extract. Using solvents with appropriate polarity significantly enhances the availability of bioactive compounds, thereby improving antimicrobial efficacy [50]. The B. minimum isolate (A2) exhibited antimicrobial activity limited to S. aureus MRSA, likely due to a lack of diverse bioactive compounds, particularly alkaloids, often linked to more substantial antimicrobial effects [51]. Although phenolic and flavonoid compounds produced by isolate A2 have been shown to possess significant antibacterial activity against S. aureus, including MRSA strains [52-54], their presence alone is insufficient to exert notable antimicrobial effects against other pathogenic microorganisms [55]. This aligns with findings in recent studies where phenolics exhibited selective activity, depending on bacterial cell wall structure and efflux pump expression [56-58].
The antibacterial activity of Bacillus sp. (A3) isolates demonstrated inhibition zones against three pathogenic microorganisms: E. coli, S. aureus, and K. pneumoniae. Previous research has indicated that Bacillus species produce various antimicrobial compounds capable of inhibiting protein synthesis [59] and disrupting bacterial cell membranes [60, 61]. The Bacillus sp. isolates exhibited a complex metabolite profile, including flavonoids, terpenoids, phenolics, and alkaloids, although the fluorescence values for alkaloids were low. The antimicrobial efficacy of these isolates against E. coli, S. aureus, and K. pneumoniae supports earlier findings that Bacillus subtilis produces active compounds effective against antibiotic-resistant bacteria [62, 63]. The presence of alkaloids in this isolate further enhances its potential to combat both gram-positive and gram-negative bacteria [61].
The B. melitensis bv isolate (A4) exhibited inhibition zones against all tested pathogenic microorganisms, indicating that its metabolites—flavonoids, terpenoids, phenolics, and alkaloids—effectively inhibit microbial growth. The presence of these compounds provides a robust foundation for the isolate's broad-spectrum antimicrobial activity, with the highest inhibition observed against C. albicans. This significant activity against C. albicans underscores the isolate's potential for treating systemic fungal infections, which are often challenging to manage with conventional antifungal agents [62]. Consequently, isolate A4 shows promise for developing novel antimicrobial agents, particularly for combination therapies targeting bacterial and fungal infections, thereby minimizing the need for multiple drugs and the associated risk of resistance [63]. Additionally, notable inhibition was also observed against P. aeruginosa. This is particularly relevant as P. aeruginosa is known for its robust intrinsic resistance mechanisms, and activity against this pathogen indicates a potent mode of action [64, 65]. Notably, there has been no prior report on the antibacterial compounds produced by B. melitensis, a bacterium traditionally recognized as a pathogen responsible for brucellosis [66, 67]. This finding is significant for its future application as a source of antibacterial compounds.
In conclusion, this study demonstrated that the bacterial isolates from Musca domestica and Chrysomya megacephala, specifically B. minimum (A2), Bacillus sp. (A3), and B. melitensis (A4), possess significant potential as sources of antimicrobial bioactive compounds. The metabolites identified—including flavonoids, terpenoids, phenolics, and alkaloids—exhibited effectiveness against antibiotic-resistant pathogens. Among the isolates, B. melitensis (A4) showed the most potent activity, with the highest inhibition zone reaching 14 mm against Acinetobacter baumannii and broad-spectrum activity against all tested pathogens.
These findings underscore the practical significance of fly-associated symbiotic bacteria as novel and underexplored sources for antimicrobial discovery. In particular, the active metabolites from B. melitensis represent promising candidates for the development of next-generation antibiotics targeting multidrug-resistant infections.
Further research is necessary to purify and identify the specific bioactive constituents within the ethyl acetate-soluble extract and elucidate the molecular mechanisms underlying their antimicrobial action. In addition, in vivo validation using animal models and subsequent clinical evaluation are warranted. The optimization of fermentation conditions and scalable biotechnological production methods is also essential to enhance metabolite yield and support future pharmaceutical applications.
We extend our gratitude to the Directorate of Research, Technology, and Community Service (DRTPM) of the Ministry of Education and Culture of the Republic of Indonesia for funding this research through the Fundamental Research Grant for the year 2024, under research contract Decision Letter No.: 0459/E5/PG.02.00/2024 and Agreement/Contract No.: 118/E5/PG.02.00.PL/2024.
[1] Grenni, P., Falconi, F. and Caracciolo, A.B. (2012). Microcosm experiments for evaluating natural bioremediation of contaminated ecosystems. Chemical Engineering Transactions, 28: 7-12. https://doi.org/10.3303/CET1228002
[2] Parasuraman, P., Pattnaik, S., Busi, S. (2019). Plant-microbe interactions in ecosystems functioning and sustainability. New and Future Developments in Microbial Biotechnology and Bioengineering: Microbial Biotechnology in Agro-Environmental Sustainability, Elsevier, pp. 255-266. https://doi.org/10.1016/B978-0-444-64191-5.00018-3
[3] Srivastava, N., Gupta, B., Gupta, S., Danquah, M.K., Sarethy, I.P. (2019). Analyzing functional microbial diversity: An overview of techniques. In Microbial Diversity in the Genomic Era, pp. 79-102. https://doi.org/10.1016/B978-0-12-814849-5.00006-X
[4] Todorov, S.D., Carneiro, K.O., Lipilkina, T.A., Do, H.K., Miotto, M., De Dea Lindner, J., Chikindas, M.L. (2024). Beneficial microorganisms for the health-promoting in oyster aquaculture: Realistic alternatives. Aquaculture International, 32: 10085-100107. https://doi.org/10.1007/s10499-024-01651-9
[5] Raja, N., Sajitha, S.T.P., Khan, M.L., G., R. (2023). Role and diversity of microbes in agriculture: Sustainable practices to promote diversity and crop productivity. Sustainable Agriculture and the Environment, Academic Press, pp. 531-555. https://doi.org/10.1016/B978-0-323-90500-8.00022-1
[6] Brits, D., Brooks, M., Villet, M.H. (2016). Diversity of Bacteria isolated from the Flies Musca domestica (Muscidae) and Chrysomya megacephala (Calliphoridae) with emphasis on vectored pathogens. African Entomology, 24(2): 365-375. https://doi.org/10.4001/003.024.0365
[7] Yin, J.H., Kelly, P.J., Wang, C. (2022). Flies as vectors and potential sentinels for bacterial pathogens and antimicrobial resistance: A review. Veterinary Sciences, 9(6): 300. https://doi.org/10.3390/VETSCI9060300
[8] Gopikrishnan, V., Radhakrishnan, M., Krupakar, P., Manigundan, K., Abirami, B., Reshma, S. (2023). Endosymbiotic interactions of actinobacteria with the insects. Microbial Symbionts: Functions and Molecular Interactions on Host, Academic Press, pp. 645-658. https://doi.org/10.1016/B978-0-323-99334-0.00001-3
[9] Grundmann, C.O., Guzman, J., Vilcinskas, A., Pupo, M.T. (2024). The insect microbiome is a vast source of bioactive small molecules. Natural Product Reports, 41(6): 935-967. https://doi.org/10.1039/D3NP00054K
[10] Koehler, S., Kaltenpoth, M. (2013). Maternal and environmental effects on symbiont-mediated antimicrobial defense. Journal of Chemical Ecology, 39: 978-988. https://doi.org/10.1007/s10886-013-0304-1
[11] Ganley, J.G., Carr, G., Ioerger, T.R., Sacchettini, J.C., Clardy, J., Derbyshire, E.R. (2018). Discovery of antimicrobial lipodepsipeptides produced by a serratia sp. within mosquito microbiomes. ChemBioChem, 19(15): 1590-1594. https://doi.org/10.1002/CBIC.201800124
[12] Kanan, M., Salaki, C., Mokosuli, Y.S. (2020). Molecular identification of bacterial species from Musca domestica L. and Chrysomya megachepala L. in Luwuk City, Central Sulawesi, Indonesia. Journal of Pure and Applied Microbiology, 14(2): 1595-1607. https://doi.org/10.22207/JPAM.14.2.58
[13] Salam, M., Bolletta, V., Meng, Y., Yakti, W., Grossule, V., Shi, D., Hayat, F. (2024). Exploring the role of the microbiome of the H. illucens (black soldier fly) for microbial synergy in optimizing black soldier fly rearing and subsequent applications. Environmental Pollution, 363: 125055. https://doi.org/10.1016/j.envpol.2024.125055
[14] Talluri, P., Lanka, S.S. (2017). Optimization of cultural parameters for the production of antimicrobial compound from Lactobacillus fermentum (MTCC No. 1745). Journal of Bacteriology & Mycology: Open Access, 4(5): 154-157. https://doi.org/10.15406/JBMOA.2017.04.00107
[15] Téfit, M.A., Gillet, B., Joncour, P., Hughes, S., Leulier, F. (2018). Stable association of a Drosophila-derived microbiota with its animal partner and the nutritional environment throughout a fly population’s life cycle. Journal of Insect Physiology, 106: 2-12. https://doi.org/10.1016/j.jinsphys.2017.09.003
[16] Matthews, M.K., Malcolm, J., Chaston, J.M. (2021). Microbiota influences fitness and timing of reproduction in the fruit fly Drosophila melanogaster. Microbiology Spectrum, 9(2): e00034-21. https://doi.org/10.1128/Spectrum.00034-21
[17] Abdel-Razek, A.S., El-Ghonemy, D.H., Shaaban, M. (2020). Production and purification of bioactive compounds with potent antimicrobial activity from a novel terrestrial fungus Aspergillus sp. DHE 4. Biocatalysis and Agricultural Biotechnology, 28: 101726. https://doi.org/10.1016/J.BCAB.2020.101726
[18] Ruiz-Sanchez, J.P., Villegas-Mendez, M.A., Montañez, J., Benavente-Valdés, J.R., Morales-Oyervides, L. (2023). Microbial production of bioactive compounds: Recent advancements and trends. Microbial Bioactive Compounds: Industrial and Agricultural Applications, Springer, Cham, pp. 1-20. https://doi.org/10.1007/978-3-031-40082-7_1
[19] Peng, Z.R., Zhang, J.G., Zhang, J.B., Lin, X.Q., Chen, W., Yang, Y.J., Liu, Z.Z. (2024). Identification and biological characteristics of Enterococcus casseliflavus TN-47 isolated from Monochamus alternatus. International Journal of Systematic and Evolutionary Microbiology, 74(4): 006305. https://doi.org/10.1099/IJSEM.0.006305
[20] Sunazuka, T., Hirose, T., Omura, S. (2008). Efficient total synthesis of novel bioactive microbial metabolites. Accounts of Chemical Research, 41(2): 302-314. https://doi.org/10.1021/AR6000044
[21] Peterson, E., Kaur, P. (2018). Antibiotic resistance mechanisms in bacteria: Relationships between resistance determinants of antibiotic producers, environmental bacteria, and clinical pathogens. Frontiers in Microbiology, 9: 426686. https://doi.org/10.3389/fmicb.2018.02928
[22] Khamesipour, F., Lankarani, K.B., Honarvar, B., Kwenti, T.E. (2018). A systematic review of human pathogens carried by the housefly (Musca domestica L.). BMC Public Health, 18: 1-15. https://doi.org/ 10.1186/s12889-018-5934-3
[23] Wiktorczyk-Kapischke, N., Skowron, K., Kwiecińska-Piróg, J., Białucha, A., Wałecka-Zacharska, E., Grudlewska-Buda, K., Kraszewska, Z., Gospodarek-Komkowska, E. (2022). Flies as a potential vector of selected alert pathogens in a hospital environment. International Journal of Environmental Health Research, 32(8): 1868-1887. https://doi.org/10.1080/09603123.2021.1919605
[24] Ghosh, S., Waite, J.L., Clayton, D.H., Adler, F.R. (2014). Can antibodies against flies alter malaria transmission in birds by changing vector behavior? Journal of Theoretical Biology, 358: 93-101. https://doi.org/10.1016/J.JTBI.2014.05.020
[25] Dawaye, D.A., Djaouda, M., Fils, E.M.B. (2021). Diversity of synanthropic flies and their potential for transmitting diarrheal diseases in Maroua (Far North- Cameroon. Pan African Medical Journal, 38: 1-15. https://doi.org/10.11604/PAMJ.2021.38.410.24687
[26] Ercan, N., Yildirim, A., Duzlu, O., Ercan, F., Yetismis, G., Pekmezci, G.Z., Inci, A. (2024). Identification and distribution of some medico-veterinary important pathogens in muscid flies in two geographical regions of Türkiye. Medical and Veterinary Entomology, 38(4): 440-448. https://doi.org/10.1111/MVE.12734
[27] Kanan, M. (2022). Antimicrobial activity of moringa leaf infusion on Escherichia coli isolate from Musca domestica L. Natural Science: Journal of Science and Technology, 11(2): 67-73. https://doi.org/10.22487/25411969.2022.V11.I02.16166
[28] Gerace, E., Mancuso, G., Midiri, A., Poidomani, S., Zummo, S., Biondo, C. (2022). Recent advances in the use of molecular methods for the diagnosis of bacterial infections. Pathogens, 11(6): 663. https://doi.org/10.3390/PATHOGENS11060663
[29] Yamin, D., Uskoković, V., Wakil, A.M., et al. (2023). Current and future technologies for the detection of antibiotic-resistant bacteria. Diagnostics, 13(20): 3246. https://doi.org/10.3390/DIAGNOSTICS13203246
[30] Cuetero-Martínez, Y., Flores-Ramírez, A., De los Cobos-Vasconcelos, D., Aguirre-Garrido, J.F., López-Vidal, Y., Noyola, A. (2023). Removal of bacterial pathogens and antibiotic resistance bacteria by anaerobic sludge digestion with thermal hydrolysis pre-treatment and alkaline stabilization post-treatment. Chemosphere, 313: 137383. https://doi.org/10.1016/J.CHEMOSPHERE.2022.137383
[31] Lin, S.W., Shelomi, M. (2024). Black soldier fly (Hermetia illucens) microbiome and microbe interactions: A scoping review. Animals, 14(22): 3183. https://doi.org/10.3390/ani14223183
[32] Gołebiowski, M., Cerkowniak, M., Urbanek, A., Dawgul, M., Kamysz, W., Boguś, M.I., Sosnowska, D., Stepnowski, P. (2014). Antimicrobial activity of untypical lipid compounds in the cuticular and internal lipids of four fly species. Journal of Applied Microbiology, 116(2): 269-287. https://doi.org/10.1111/JAM.12370
[33] Chernysh, S., Gordya, N., Suborova, T. (2015). Insect antimicrobial peptide complexes prevent resistance development in bacteria. PLOS One, 10(7): e0130788. https://doi.org/10.1371/JOURNAL.PONE.0130788
[34] Bouaziz, A., Mhalla, D., Zouari, I., Jlaiel, L., Tounsi, S., Jarraya, R., Trigui, M. (2016). Antibacterial and antioxidant activities of Hammada scoparia extracts and its major purified alkaloids. South African Journal of Botany, 105: 89-96. https://doi.org/10.1016/J.SAJB.2016.03.012
[35] Martínez-Fructuoso, L., Arends, S.J.R., Freire, V.F., et al. (2023). Screen for new antimicrobial natural products from the NCI Program for Natural Product Discovery Prefractionated Extract Library. ACS Infectious Diseases, American Chemical Society. 9(6): 1245-1256. https://doi.org/10.1021/acsinfecdis.3c00067
[36] Bouymajane, A., Filali, F.R., Moujane, S., Majdoub, Y.O. El, Otzen, P., Channaoui, S., Ed-Dra, A., Bouddine, T., Sellam, K., Boughrous, A.A., Miceli, N., Altemimi, A.B., Cacciola, F. (2024). Phenolic compound, antioxidant, antibacterial, and in silico studies of extracts from the aerial parts of Lactuca saligna L. Molecules, 29(3): 596. https://doi.org/10.3390/MOLECULES29030596
[37] Rahman, M.M., Rahaman, M.S., Islam, M.R., Rahman, F., Mithi, F.M., Alqahtani, T., Almikhlafi, M.A., Alghamdi, S.Q., Alruwaili, A.S., Hossain, M.S., Ahmed, M., Das, R., Emran, T.B., Uddin, M.S. (2021). Role of phenolic compounds in human disease: Current knowledge and future prospects. Molecules, 27(1): 233. https://doi.org/10.3390/MOLECULES27010233
[38] Chagas, M.S. do S., Behrens, M.D., Moragas-Tellis, C.J., Penedo, G.X.M., Silva, A.R., Gonçalves-De-Albuquerque, C.F. (2022). Flavonols and flavones as potential anti-inflammatory, antioxidant, and antibacterial compounds. Oxidative Medicine and Cellular Longevity, 2022(1): 9966750. https://doi.org/10.1155/2022/9966750
[39] Mehmood, A., Javid, S., Khan, M.F., Ahmad, K.S., Mustafa, A. (2022). In vitro total phenolics, total flavonoids, antioxidant and antibacterial activities of selected medicinal plants using different solvent systems. BMC Chemistry, 16: 1-10. https://doi.org/10.1186/s13065-022-00858-2
[40] Donadio, G., Mensitieri, F., Santoro, V., Parisi, V., Bellone, M.L., De Tommasi, N., Izzo, V., Dal Piaz, F. (2021). Interactions with microbial proteins driving the antibacterial activity of flavonoids. Pharmaceutics, 13(5): 660. https://doi.org/10.3390/pharmaceutics13050660
[41] Huang, W., Wang, Y., Tian, W., Cui, X., Tu, P., Li, J., Shi, S., Liu, X. (2022). Biosynthesis investigations of terpenoid, alkaloid, and flavonoid antimicrobial agents derived from medicinal plants. Antibiotics, 11(10): 1380. https://doi.org/10.3390/antibiotics11101380
[42] Dousti, B., Nabipor, F. (2021). Evaluation of the phenolic contents and antibacterial activity of different concentrations of Onosma chlorotricum Boiss. Biotechnologia, 102(4): 377-386. https://doi.org/10.5114/BTA.2021.111095
[43] Othman, L., Sleiman, A., Abdel-Massih, R.M. (2019) Antimicrobial activity of polyphenols and alkaloids in middle eastern plants. Frontiers in Microbiology, 10: 911. https://doi.org/10.3389/FMICB.2019.00911
[44] Wang, C., Du, W., Lu, H., Lan, J., Liang, K., Cao, S. (2021). A review: Halogenated compounds from marine actinomycetes. Molecules, 26(9): 2754. https://doi.org/10.3390/molecules26092754
[45] Heo, C.S., Kang, J.S., Yang, J.W., Lee, M.A., Lee, H.S., Shin, H.J. (2024). Labdane-type diterpenoids from Streptomyces griseorubens and their antimicrobial and cytotoxic activities. International Journal of Molecular Sciences, 25(6): 3311. https://doi.org/10.3390/ijms25063311
[46] Zhao, M.Y., Shi, X.B., Chang, J.H., Wang, R.X., Zhou, J.Y., Liu, P. (2025). Amorphous solid dispersions of glycyrrhetinic acid: Using soluplus, PVP, and PVPVA as the polymer matrix to enhance solubility, bioavailability, and stability. AAPS PharmSciTech, 26: 18. https://doi.org/10.1208/s12249-024-03007-1
[47] Isreb, A., Alhnan, M.A., Mkia, A., Al-Jammal, K., Yaghi, A., Oga, E.F., Timmins, P., Bonner, M., Forbes, R.T. (2025). Evaluation of drug–polymer and drug–drug interaction in cellulosic multi-drug delivery matrices. Methods and Protocols, 8(1): 4. https://doi.org/10.3390/mps8010004
[48] Noviany, Sialdian, D., Setiawan, A., Irawan, B., Azmi, M.N., Hadi, S. (2021). Bioassay-guided separation approach for characterization of new antibacterial fractions from the stem roots extracts of archidendron jiringa. Journal of the Turkish Chemical Society, Section A: Chemistry, 8(2): 391-402. https://doi.org/10.18596/jotcsa.831054
[49] Wang, F., Cheng, L., Cao, Y., Wei, Q., Tong, C., Shi, S. (2022). Online extraction and enrichment coupling with high-speed counter-current chromatography for effective and target isolation of antitumor anthraquinones from seeds of Cassia obtusifolia. Journal of Separation Science, 45(4): 938-944. https://doi.org/10.1002/jssc.202100775
[50] Guo, L., Sun, Y., Ping, X., Liu, J., Wang, X., Qin, N. (2022). Chemical composition and antibacterial activity of ethyl acetate extract of Astragalus membranaceus aerial parts. Journal of Food Safety, 42: e12947. https://doi.org/10.1111/JFS.12947
[51] Du, H., Wang, Y., Hao, X., Li, C., Peng, Y., Wang, J., Liu, H., Zhou, L. (2009). Antimicrobial phenolic compounds from Anabasis Aphylla L. Natural Product Communications, 4(3): 385-388. https://doi.org/10.1177/1934578X0900400314
[52] Li, C., Yan, W., Cui, E., Zheng, C. (2021). Anti-bacterial effect of phytoconstituents isolated from Alimatis rhizoma. Applied Biological Chemistry, 64: 1-5. https://doi.org/10.1186/s13765-020-00583-1
[53] Bouarab-Chibane, L., Forquet, V., Lantéri, P., Clément, Y., Léonard-Akkari, L., Oulahal, N., Degraeve, P., Bordes, C. (2019). Antibacterial properties of polyphenols: Characterization and QSAR (Quantitative structure-activity relationship) models. Frontiers in Microbiology, 10: 440698. https://doi.org/10.3389/FMICB.2019.00829
[54] Kauffmann, A.C., Castro, V.S. (2023). Phenolic compounds in bacterial inactivation: A perspective from Brazil. Antibiotics, 12(4): 645. https://doi.org/10.3390/ANTIBIOTICS12040645
[55] Veiko, A.G., Olchowik-Grabarek, E., Sekowski, S., Roszkowska, A., Lapshina, E.A., Dobrzynska, I., Zamaraeva, M., Zavodnik, I.B. (2023). Antimicrobial activity of quercetin, naringenin and catechin: Flavonoids inhibit Staphylococcus aureus-induced hemolysis and modify membranes of bacteria and erythrocytes. Molecules, 28(3): 1252. https://doi.org/10.3390/MOLECULES28031252
[56] Lo, M.M., Benfodda, Z., Dunyach-Rémy, C., Bénimélis, D., Roulard, R., Fontaine, J.X., Mathiron, D., Quéro, A., Molinié, R., Meffre, P. (2022). Isolation and identification of flavones responsible for the antibacterial activities of Tillandsia bergeri extracts. ACS Omega, 7(40): 35851-35862. https://doi.org/10.1021/acsomega.2c04195
[57] Hellewell, L., Bhakta, S. (2020). Chalcones, stilbenes and ketones have anti-infective properties via inhibition of bacterial drug-efflux and consequential synergism with antimicrobial agents. Access Microbiology, 2(4): 1-12. https://doi.org/10.1099/acmi.0.000105
[58] Moreno-Chamba, B., Salazar-Bermeo, J., Navarro-Simarro, P., Narváez-Asensio, M., Martínez-Madrid, M.C., Saura, D., Martí, N., Valero, M. (2023). Autoinducers modulation as a potential anti-virulence target of bacteria by phenolic compounds. International Journal of Antimicrobial Agents, 62(4): 106937. https://doi.org/10.1016/j.ijantimicag.2023.106937
[59] Macêdo, N.S., De Sousa Silveira, Z., Cordeiro, P.P.M., et al. (2022). Inhibition of Staphylococcus aureus efflux pump by O-eugenol and its toxicity in drosophila melanogaster animal model. BioMed Research International, 2022: 440996. https://doi.org/10.1155/2022/1440996
[60] Wu, D., Fu, L., Cao, Y., Dong, N., Li, D. (2023). Genomic insights into antimicrobial potential and optimization of fermentation conditions of pig-derived Bacillus subtilis BS21. Frontiers in Microbiology, 14: 1239837. https://doi.org/10.3389/fmicb.2023.1239837
[61] Abriouel, H., Franz, C.M.A.P., Omar, N.B., Gálvez, A. (2011). Diversity and applications of Bacillus bacteriocins. FEMS Microbiology Reviews, Oxford Academic. 35(1): 201-232. https://doi.org/10.1111/J.1574-6976.2010.00244.X
[62] Sumi, C.D., Yang, B.W., Yeo, I.C., Hahm, Y.T. (2015). Antimicrobial peptides of the genus Bacillus: A new era for antibiotics. Canadian Journal of Microbiology, 61(2): 93-103. https://doi.org/10.1139/CJM-2014-0613
[63] de Souza Freitas, F., Coelho de Assis Lage, T., Ayupe, B.A.L., de Paula Siqueira, T., de Barros, M. and Tótola, M.R. (2020). Bacillus subtilis TR47II as a source of bioactive lipopeptides against Gram-negative pathogens causing nosocomial infections. 3 Biotech, 10: 1-10. https://doi.org/10.1007/S13205-020-02459-Z
[64] Medeot, D.B., Fernandez, M., Morales, G.M., Jofré, E. (2020). Fengycins from Bacillus amyloliquefaciens MEP218 exhibit antibacterial activity by producing alterations on the cell surface of the pathogens Xanthomonas axonopodis pv. vesicatoria and Pseudomonas aeruginosa PA01. Frontiers in Microbiology, 10: 1-12. https://doi.org/10.3389/FMICB.2019.03107
[65] Yang, J., Xu, J.F., Liang, S. (2024). Antibiotic resistance in Pseudomonas aeruginosa: Mechanisms and emerging treatment. Critical Reviews in Microbiology. https://doi.org/10.1080/1040841X.2024.2429599
[66] Jeong, G.J., Khan, F., Tabassum, N., Kim, Y.M. (2024). Natural and synthetic molecules with potential to enhance biofilm formation and virulence properties in Pseudomonas aeruginosa. Critical Reviews in Microbiology, 50(5): 830-858. https://doi.org/10.1080/1040841X.2023.2282459
[67] Bartal, A., Huynh, T., Kecskeméti, A., Vörös, M., Kedves, O., Allaga, H., Varga, M., Kredics, L., Vágvölgyi, C., Szekeres, A. (2023). Identifications of surfactin-type biosurfactants produced by bacillus species isolated from rhizosphere of vegetables. Molecules, 28(3): 1172. https://doi.org/10.3390/MOLECULES28031172