Efficacy of Trichoderma asperellum and Pseudomonas aeroginosa Metabolites Against Fusarium Wilt in Bananas

Alfizar Alfizar*![]() | Amda Resdiar
| Amda Resdiar![]() | Iwandikasyah Putra
| Iwandikasyah Putra![]() | Hendri Bustamam
| Hendri Bustamam![]() | Mohammad Adzannie Bessania
| Mohammad Adzannie Bessania![]() | Syaukani Syaukani
| Syaukani Syaukani![]()
© 2023 IIETA. This article is published by IIETA and is licensed under the CC BY 4.0 license (http://creativecommons.org/licenses/by/4.0/).
OPEN ACCESS
Banana cultivation is increasingly compromised by pests and diseases, notably the devastating impact of Fusarium oxysporum f.sp. cubense (Foc) wilt disease in Indonesian plantations. This study was designed to characterize and identify biological agents and to evaluate the effectiveness of their metabolites against Foc wilt. A randomized block design was employed, comprising four treatments with six replications each (n=24). The treatments included: distilled water as a control (T0); metabolites derived from Trichoderma asperellum (T1); metabolites from Pseudomonas aeruginosa (T2); and a combination of metabolites from both T. asperellum and P. aeruginosa (T3). Molecular identification confirmed the agents as T. asperellum and P. aeruginosa, with sequence homology of 100% and 98.46%, respectively. The control group's incubation period spanned 281.52 days, during which 100% disease incidence was recorded. Conversely, treatments utilizing the secondary metabolites of T. asperellum, P. aeruginosa, and their combination exhibited no symptomatology up to the 10-month flowering period. The efficacy of the secondary metabolites in inhibiting Foc infection was thus demonstrated up to the critical flowering stage. Results indicate that both the individual and combined applications of T. asperellum and P. aeruginosa metabolites are effective in suppressing Fusarium wilt in bananas. The integration of these biological agents into disease management strategies offers a promising avenue for mitigating the impact of Foc on banana agriculture.
banana, fusarium, trichoderma asperellum, pseudomonas aeroginosa, metabolites
Bananas are highly esteemed in Indonesia for their palatability, their nutritional profile—rich in potassium, fiber, protein, and vitamins—and their versatility in culinary applications, ranging from fresh consumption to their use in an array of processed foods and beverages [1-3]. The country's banana industry is robust, with national production figures for 2020 reported at over 8 million tons, and the Aceh Province alone contributing 65,366 tons, thus emerging as a leading producer [4, 5]. However, the sustainability of banana cultivation is imperiled by biotic stressors, notably the Fusarium oxysporum f.sp. cubense (Foc) wilt disease, which has wrought havoc from Aceh to Papua [6]. The soil-borne pathogen Fusarium oxysporum, particularly the Tropical Race 4 (TR4) or Fusarium odoratissimum species, is identified as the causative agent of the disease, with its prevalence in fields correlating positively with disease incidence and severity [7-9]. This pathogen is infamous for the historical obliteration of the 'Gros Michel' variety in Panama, illustrating the potentially catastrophic consequences of Fusarium wilt, or Panama disease [10].
The control of soil pathogens through fungicides is constrained by the interaction of these chemicals with soil organic matter, which often renders them ineffective [11]. Furthermore, the indiscriminate use of fungicidal compounds is associated with toxic residues that detrimentally impact soil biota, thereby challenging the environmental sustainability of such practices [12]. In this context, secondary metabolites emerge as a viable alternative, offering promise for pest and disease management in agricultural settings [13, 14]. Among these, the genus Trichoderma has garnered attention for its antagonistic potential against various phytopathogens. For instance, Trichoderma harzianum has been shown to produce secondary metabolites that are efficacious in controlling Fusarium oxysporum f.sp. lycopersici in tomatoes [15]. Trichoderma species not only serve as biocontrol agents but also play a role in soil organic matter decomposition. In vitro studies have demonstrated that Trichoderma sp. can antagonize Foc, inhibiting its colony growth by up to 86% through mechanisms such as hyperparasitism [16]. Additionally, Trichoderma virens has been reported to produce indole-3-acetic acid (IAA) and to inhibit Ganoderma boninense in palm oil nurseries [17]. Its efficacy extends to controlling Colletotrichum gloeosporioides in chili [18], suppressing Alternaria porri in onions [19], and exerting antibiotic properties as a biocontrol agent [20].
Bacteria, particularly strains of Pseudomonas spp., have also been recognized for their biocontrol capabilities. Pseudomonas aeruginosa, for example, is noted for its production of phytohormones, phosphate solubilization, and antagonism against soil-borne pathogens [21, 22]. Certain strains of Pseudomonas spp. have proven effective in field control of Foc, underscoring their potential as biological control agents against Fusarium Wilt of Banana (FWB) [23].
The fungal isolate employed in this study was sourced from disease-free banana tubers at the University Farm of Universitas Teuku Umar (UTU), Indonesia, while the bacterial isolate was obtained from the Phytopathology Laboratory, Faculty of Agriculture, Universitas Bengkulu, Indonesia. Both cultures were maintained at 4 degrees Celsius and re-cultured for freshness prior to utilization. The objective of this study is twofold: to characterize and identify the biological agents, and to evaluate the efficacy of their secondary metabolites against Fusarium wilt disease in bananas.
2.1 Isolation of microbes as a biological agent
This study utilizes fungi and bacteria as biological agents and microorganisms. Fungus isolates were cultured using Potato Dextrose Agar (PDA) medium (4g/L potato extract, 20g/L glucose, 15 g/L agar, Oxoid™) while bacterial isolates were cultured on King's B medium (20g/L protease peptone no. 3, 1.5 g/L K2HPO4, 1,5 g/L MgSO4.7H2O, 15 g/L agar, 10 ml glycerol, HiMedia®).
2.2 Morphological characterization of biological agent microorganisms
The characterization of bacterial and fungal isolates involved macroscopic and microscopic observations. We conducted this research in the Plant Pathology Laboratory, Faculty of Agriculture, Universitas Syiah Kuala. The morphological characterization of bacterial isolates included the statement of colony form, color, margin, elevation, and Gram staining for microscopic examination. Similarly, the morphological characteristics of fungal isolates were assessed by macroscopic examination, which involved observing the colony shape and color, as well as microscopic examination to determine the morphology of hyphae, whether they were septate or non-septate, and the condition of conidia.
2.3 Molecular identification of biological agent microorganisms
2.3.1 Primers
The molecular identification was conducted at Genetika Science Ind. Laboratory, Banten, Indonesia. The primary sequence used in this research is the 16S rRNA region marker, utilized for bacterial identification, and ITS1-ITS4, a general primer that amplifies the internal transcribed spacer region for purpose identification.
2.3.2 DNA isolation
DNA of microorganism was extracted using the Quick-DNA Fungal/Bacterial Miniprep Kit (Zymo Research, D6005) per the manufacturer's procedure recommended.
2.3.3 DNA amplification
PCR analysis was performed in two reactions, the first for 16S Primer and the second for ITS1-ITS4 primer. PCR amplification with specific primer sets was carried out using the MyTaq HS Red Mix, 2X (Bioline, BIO-25048). The amplification of the 16S gene region, the PCR reaction mixture was prepared as follows: in a total volume of 25 µl, consisting of 9.5 µl of ddH2O, 12.5 µl of MyTaq HS Red Mix, 2X, 1 µl of 10 µM 27F primer (Table 1), and 1 µl of DNA template.
Table 1. The primary sequences were used in the study
|
PCR Reaction |
Primer Sequences |
|
16S (27F/1492R) PRIMER |
F: AGAGTTTGATCMTGGCTCAG |
|
R: GGTTACCTTGTTACGACTT |
|
|
ITS1-ITS4 PRIMER |
F: TCCGTAGGTGAACCTGCGG |
|
R: TCCTCCGCTTATTGATATGC |
The amplification of the ITS1-ITS4 region, the PCR reaction mixture contained 9.5 µl of ddH2O, 12.5 µl of MyTaq HS Red Mix, 2X, 1 µl of 10 µM ITS1 primer (Table 1), 1 µl of 10 µM ITS4 primer (Table 1), and 1 µl of DNA template, in a total volume of 25 µl. The PCR for both reactions was run as follows: initial denaturation at 95℃ for 3 minutes, followed by 35 cycles of denaturation at 95℃ for 15 seconds, annealing at 52℃ for 30 seconds, extension at 72℃ for 45 seconds, and a final extension step at 72℃ for 3 minutes.
PCR product (5 µl of DNA template combined with 2 µl of loading dye) were separated by agarose gel electrophoresis (1.2%), stained with ethidium bromide solution (0.01%), 47 minutes in 50 volts, and detected with UV transluminator.
PCR product (1µl) was separated by agarose gel electrophoresis (0.8%), stained with ethidium bromide solution (0.01%), 47 minutes in 50 volts, and visualization with UV transluminator. Bidirectional sequencing was performed to combine the top and bottom strands of the initial double-stranded DNA fragment in separate reactions using BIOEDIT software. The BLASTN program was used to compare the sequence to examine the similarity. Related analysis with phylogenetic trees using the neighbor-joining method was performed by aligning the sample sequences with the NCBI database using the Molecular Evolutionary Genetics Analysis (MEGA 11.0.13) software (New York, USA). Relatedness analysis with phylogenetic tres was performed [24].
2.4 Isolation of Fusarium oxysporum
The Fusarium oxysporum strain with pathogenic properties was isolated from a symptomatic banana tuber showing typical signs of Fusarium wilt infection. The tuber was obtained from the banana collection garden at the University Farm of Teuku Umar University in Indonesia. The tuber was thoroughly cleansed and rinsed to remove soil pollutants to achieve aseptic conditions. The symptomatic part was then divided into pieces that were 1cm × 1cm × 1cm in size. Surface sterilization of these fragments was performed by treating them with a 5% chlorine solution for 1 minute, followed by immersion in alcohol for 1 minute, and finally, rinsing twice with sterile distilled water. Following sterilization, the fragments were placed on a (PDA) medium supplemented with Streptomycin at 50 mg.l-1. The cultures were incubated in a temperature-controlled incubator set at 27℃. As the fungal growth appeared, we transferred it to a fresh PDA medium to ensure the purity of the culture. We repeated this process until pure cultures of the Fusarium strain were obtained, free from contamination and other microorganism.
2.5 Production of secondary metabolites of bacterial and fungal isolates
Pure cultures of bacteria and fungi were obtained and cultured in liquid media to promote the production of secondary metabolites. For the fungal isolate, the liquid medium consisted of a combination of coconut water, rice water, and sugar. The propagation process involved meticulously blending 4 liters of rice washing water, 1 liter of coconut water, and 75 grams of sugar. The resulting mixture was thoroughly mixed, sterilized in an autoclave at 121℃ for 30 minutes, cooled down, and transferred into a sterile jerry can. A pure culture of the fungal isolate was diluted with 10 ml of clean distilled water and added to the jerry can, which was sealed tightly to initiate propagation. The jerry can then be placed on a shaker operating at 150 rpm for ten days, providing optimal conditions for metabolite production. In the case of the bacterial isolate, a liquid medium consisting of golden snail broth supplemented with shrimp paste and sugar was utilized for secondary metabolite propagation. The process involved obtaining 400 grams of gold snail meat and combining it with 2 grams of shrimp paste and 500 ml of distilled.
The golden snail meat mixture and shrimp paste were boiled until the resulting broth was carefully filtered and transferred to a sterile jerry can. A pure culture of the bacterial isolate was suspended in 10 ml of distilled water and added to the jerry can, which was then securely sealed. The jerry can be placed on a shaker operating at 15 rpm for ten days, allowing for optimal growth and metabolite production by the bacterial isolate. Both were filtered with a polycarbonate membrane (0.2 m, 25 mm diameter, millipore, PC 110656N) to get pure secondary metabolites by removing the mycelium of fungus and bacterium cells.
2.6 Treatment of bacterial and fungal secondary metabolites as a biocontrol method
The contour of the experimental field is flat, located in the campus area, and has never been used for research. A fence surrounded the research plot to avoid animals. The field was cleared of grass, and a planting hole was made according to planting distance. The design used in this study was a randomized block design with four treatments and six replications, with 24 units of experimental. The determination of the experimental design is based on the small number of treatments. Each treatment unit consisted of 4 banana seedlings aged two months, with 96 banana seedlings. Analysis of variance one way was carried out using the statistical analysis system IBM SPSS version 24 program to test the effect of treatment on the observed variables. Then, each treatment that had a significant effect was carried out by LSD test to see the differences in each treatment at the 5% level. The variety of banana (Musa acuminata) seeds used were two months old and taken from healthy banana clumps at the University Farm, Teuku Umar University. The method of treating the tuber of banana seedlings was as follows. The banana tubers and roots were cleaned from the soil, washed, and then soaked in a suspension of secondary metabolites according to the experimental treatment for 12 hours.
The treatments were as follows: they were soaked in distilled water as control (T0), in a suspension of T. asperellum metabolites (T1), in a suspension of P. aeroginosa metabolites (T2), and in a rest of a mixture of T. asperellum and P. aeroginosa metabolites (T3). Before planting in the field, 20 ml containing 106.ml-1 suspension of Fusarium spores was sprayed evenly into all planting holes using a hand sprayer. Then, banana seedlings that had received each treatment were planted in planting holes with a spacing of 2 × 2 meters. The parameter observed was the incubation period (days). The incubation period was monitored every day from the first day after treatment until the initial symptoms appeared. If the leaves showed yellowing signs, the initial symptoms could be counted. The formula calculated the Incidence Parameters of the disease:
$I P=\frac{n}{N} \times(100) \%$
where, n is the number of infected plants, and N is the number of plants observed. Parameters of disease severity were measured by looking at the tuber's discoloration and leaves wilting. The results of observations/scores were calculated using the formula:
$K P=\frac{\sum_{i=0}^i\left(n_i \times v_i\right)}{N \times V} \times 100 \%$
where, ni is the first score, the number of infected plants; vi is the value of the I score; N is the number of plants observed; and V is the highest score in the scoring reference.
The disease severity (LSI = leaf symptom index) was carried out based on the symptoms of wilting on the leaves, and the severity of discoloration in the tuber vascular tissue (RDI = rhizome discoloration index) was carried out based on the percentage of discoloration on the tuber that was split horizontally. The scores and categories used in the LSI measurement, namely: score 0 with the percentage of leaf area symptomatic area (X) being 0 or no yellowing symptoms; score 1 with 1 X < 25 if the lower leaf edges begin to turn yellow; score 2 with 26 X < 50 if all the lower leaves turn yellow; score 3 with 51 X < 75 if all yellow banana leaves are found; and score 4 with 76 X < 100 if the plant dies. The categories used to measure the RDI are as follows: score 0 if the percentage of symptom area in the tuber (Y) is 0 or there are no symptoms of necrosis in the tuber tissue; score 1 with 1 Y < 25 if there is a little necrosis in the tissue in the tuber; score 2 with 26 Y < 50 if there is quite a lot of necrosis in the tuber tissue; score 3 with 51 Y < 75 if most of the necrotic tuber tissue is found; and score 4 with 76 X < 100 if all necrotic tuber tissue is found.
2.7 Data analysis
The data were analyzed using the IBM SPSS Version 25 program, and if the analysis of variance showed significant differences, then further tested using LSD 5%. DNA sequencing was performed, trimmed, and assembled with the BioEdit program 7.2.5 (Denton, USA) and then converted into FASTA form. Furthermore, the result was analyzed using the Basic Local Alignment Search Tool (BLAST) program on the NCBI website (https://www.ncbi.nlm.nih.gov/) to determine the identity of the reference sequences contained in GeneBank.2.7 Kinship Analysis. Bacterial and fungal relationships were determined using phylogenetic analysis, which they performed by aligning the analyzed DNA sequences with homologous samples downloaded from GeneBank. The phylogenetic tree was then constructed based on the 16S rRNA (bacteria) and ITS (fungi) gene sequence using the Molecular Evolutionary Genetics Analysis (MEGA 11.0.13) software (New York, USA).
3.1 Morphological characterization of biological agent microorganism
The fungal isolate observed distinct morphological characteristics, with the colony initially appearing white and gradually transitioning to green, eventually becoming dark green over time. The conidia were spherical (globose) in shape, green in color, and exhibited numerous branching conidiophores. Notably, the presence of short stalks on the fields was observed, indicating its classification as Trichoderma sp (Figure 1). Conversely, the bacterial isolate exhibited specific morphological characteristics. The bacterial colonies appeared circular, with a smooth surface and a flat elevation. They displayed a creamy yellowish color and were relatively small in size. Microscopic examination revealed that these bacteria were Gram-positive, with a basil-like shape, suggesting their identification as Pseudomonas aeruginosa (Figure 2).
Figure 1. Morphological characteristics of Trichoderma sp. (a) Colony morphology, (b) Conidia under microscope
Figure 2. Morphological characteristics of Pseudomonas aeruginosa (a) Colony morphology, (b) Gram-positive bacilli cell under a microscope with 1000x magnification
Molecular identification of biological agent microorganisms The morphological characteristics of fungal isolate a colony color that was initially white, then turned green, and over time it is dark green; the conidia are round (globose) and the shade of green; there are many branching conidiophores; the short fialid stalk is characteristic; which indicated as Trichoderma sp.
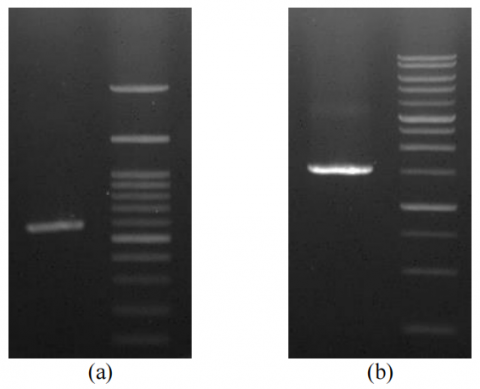
The results of PCR showed only a single band with a size of 597 bp for fungi isolate and 1427 bp for bacteria isolate, which indicates that their ITS and 16S rRNA gene have been well amplified (Figure 3). DNA can be visualized by ultraviolet (UV) because the agarose gel contains ethidium bromide, whose molecule fluoresces when illuminated with UV light in the visible UV range. Ethidium binds to the hydrophobic location of the DNA molecule by inserting it between the base bonds in the double-stranded DNA through Van der Walls interactions. The ITS gene sequencing of fungi isolates and 16S rRNA gene sequencing of the bacteria isolated in the form of an electropherogram was then translated using the BIOEDIT software to produce nucleotide sequence data for the 16S rRNA gene isolate (Figure 4).
Figure 3. (a) Gel photo of a single band with a size of 597 bp for fungi isolate with 1 µL PCR Products was assessed by electrophoresis with 1% TBE agarose, (b) 1427 bp for bacteria isolate with 1 µL PCR Products were assessed by electrophoresis with 0.8% TBE agarose
Figure 4. (a) The ITS gene sequencing of fungi isolates, (b) 16S rRNA gene sequencing of the bacteria isolate
Percentage identity shows a similarity between query sequence and target sequence. For microorganisms, using 16srRNA primers, similar to species level (above 97.5%), and for genus (above 95%) [25].
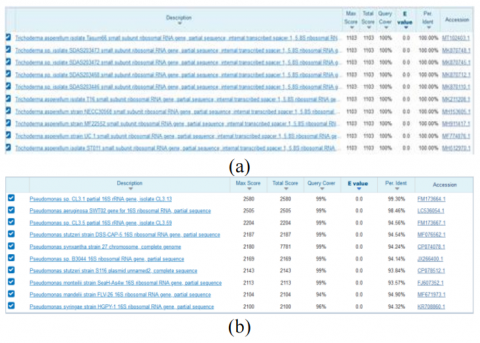
Based on the top 10 hit BLAST results against the NCBI database, the percentage identity of the fungus of Trichoderma asperellum was 100 percent, while for bacteria of Pseudomonas aeroginosa was 98,46 percent (Figure 5).
Figure 5. Percentage identity and similarity of fungus isolates (a) and bacteria isolate (b)
Further, sequencing was performed to sequence the initial double-stranded DNA fragment in separate reactions. The resulting sequences were then compared using the BLASTN. They aligned the simple sequences with the NCBI database using the Molecular Evolutionary Genetic Analysis (MEGA 11.0.13).
A phylogenetic tree (Figure 6) is a diagram that represents evolutionary relationships among organisms. Based on the analysis of the BLAST-N program, we found that fungal isolates had a 100% identity with the Trichoderma asperellum isolate Tasum66, Trichoderma asperellum isolate SDAS203473, and Trichoderma asperellum isolates SDAS203472 (Figure 6a). Meanwhile, bacterial isolates had a 99.93% identity with Pseudomonas sp. isolate CL3.13 and 98.46% identity with Pseudomonas aeruginosa (Figure 6b). Isolates with a 16S rRNA sequence homology level of more than 97% can represent the same species level. In comparison, those between 93-97% denote the identity of bacteria at the genus at the varying species level. If the homology level is below 93%, it is a new species whose nitrogen base sequence has not been included in the GenBank database. The fungal and bacterial gene sequencing obtained was made into a phylogenetic tree to determine the level of their kinship. The phylogenetic tree construction aims to visualize the relationship between sample organisms based on their evolutionary relationships with comparison organism sequences from National Center for Biotechnology Information (NCBI) sites.
Figure 6. Phylogenetic tree of fungi isolates identified as Trichoderma asperellum (a), and bacterial isolates identified as Pseudomonas aeroginosa (b)
3.2 Treatment of secondary metabolites of fungi and bacteria as biocontrol
The findings of this study, as determined by the F test of 5% (Table 2), revealed significant differences among the treatments involving secondary metabolites of Trichoderma asperellum, Pseudomonas aeruginosa, and a combination of both when compared to the control treatment across all assessed parameters. At the 5% LSD value, the confidence intervals in the control treatment (T0) were 0.567 for the incubation period, 0.00 for disease incidence, 0.701 for tuber discoloration, and 0.494 for discoloration of the leaf. In contrast, the confidence intervals were 0 in the other treatments. This research shows that it was implemented well and had high accuracy. During the research, it was seen that only the control treatment was affected by Fusarium wilt disease, while other treatments were not affected. The control treatment exhibited an incubation period of 281.52 days after planting, significantly different from the treatments involving secondary metabolites. We observed that the Fusarium sp pathogen required a prolonged incubation to infect the banana roots and tubers and subsequently manifest symptoms. In contrast, the treatments involving secondary metabolites of Trichoderma asperellum, Pseudomonas aeruginosa, and the combination of both showed no signs.
Table 2. Incubation Period, disease incidence, and disease severity of tuber discoloration and leaf wilting due to application of secondary metabolites
|
Treatment |
Incubation Period (**) |
Disease Incidence (**) |
Disease Severity (**) |
|
|
Tuber Discoloration |
Leaf Wilting |
|||
|
|
(days) |
(%) |
(%) |
(%) |
|
T0 |
281.52 b |
100 b |
72.57 b |
75.23 b |
|
T1 |
0.00 a |
0.00 a |
0.00 a |
0.00 a |
|
T2 |
0.00 a |
0.00 a |
0.00 a |
0.00 a |
|
T3 |
0.00 a |
0.00 a |
0.00 a |
0.00 a |
*Numbers followed by different letters indicate a significant difference in the LSD test at α 5%
Regarding disease incidence, the control treatment had the highest percentage, reaching 100%, and this was significantly different from the other treatments. Assessment of disease severity demonstrated that the control treatment exhibited a necrosis percentage of 72.57% in the tubers and 75.23% in leaf wilting. Conversely, the other treatments did not experience disease severity, indicating the efficacy of secondary metabolites from Trichoderma asperellum and Pseudomonas aeruginosa in suppressing the development of the Fusarium sp pathogen.
Numerous researchers have demonstrated the efficacy of Trichoderma asperellum and Pseudomonas aeruginosa in controlling various plant diseases, particularly those caused by soil-borne pathogens [26] and demonstrated the effectiveness of T. asperellum B1902 in inhibiting the growth of Fusarium oxysporum f. sp. under in vitro and in vivo conditions. Trichoderma asperellum TKD was reported to suppress the growth of F. oxysporum f.sp. capsici, resulting in improved plant height, increased fresh leaf count, and reduced dry leaf count [27]. Trichoderma sp. and Pseudomonas fluorescens were found to produce antibiotics, enzymes, toxins, and hormones that promote better plant growth [28]. Besides that, the Pseudomonas aeruginosa can control fusarium wilt disease and enhance the development of shallot plants [29]. Pseudomonas aeruginosa, an antagonistic bacterium, produces siderophores and volatile compounds such as ammonia, alkyl pyrones, and hydrogen cyanide (HCN) that are toxic to other plant pathogens, particularly F. oxysporum. Siderophores and HCN play a crucial role in the biocontrol activity against fungal pathogens as they sequester the necessary Fe ions required by F. oxysporum for germination [30]. Chitinolytic bacteria, including Pseudomonas aeruginosa found in the rhizosphere area, exhibit chitinase activity, which can induce abnormalities in the hyphae of the F. oxysporum pathogen by releasing chitinase enzymes [31].
Figure 7. (a) Discoloration on the banana tuber, (b) Leaf wilting symptom
Additionally, secondary metabolites of Trichoderma sp. have shown potential in inhibiting Pseudomonas solanacearum wilt disease in potatoes [32]. These metabolites have also effectively reduced the severity of vascular streak dieback (VSD) in cocoa plants [33]. Overall, secondary metabolites derived from Trichoderma sp. and Pseudomonas aeruginosa present promising prospects for preventing soil-borne diseases that are challenging to detect. Nevertheless, further long-term research is necessary to obtain comprehensive insights and confirm the efficacy of secondary metabolites across multiple growth periods of banana tillers in field conditions.
The effectiveness of secondary metabolites derived from Trichoderma asperellum, Pseudomonas aeruginosa, and their combination demonstrated satisfactory outcomes throughout the growth of the banana plants until they began bearing fruits at ten months of age. The 12-hour immersion process allowed the secondary metabolites to be absorbed by the roots and tubers of the banana seeds. Notably, the secondary metabolites of Trichoderma asperellum contain antibiotics, enzymes, toxins, and growth regulators that can inhibit and even eliminate Fusarium oxysporum, the causative agent of wilt disease in bananas. The control treatment appeared more susceptible to Fusarium oxysporum infection due to the absence of protective substances in the banana roots and tubers. Consequently, the Fusarium oxysporum pathogen within the planting holes had sufficient access to nutrients from exudates and other plant metabolic products released in the rhizosphere. This favorable environment and the absence of protective materials allowed the dynamic development of the fusarium, leading to early infection of the banana roots and tubers, then the symptoms appear on the leaves (Figure7b).
Consequently, symptoms began to emerge around nine months of banana plant growth. The incubation period for Fusarium oxysporum was observed to be long, approximately 281.52 days or over nine months. Upon splitting a symptomatic banana plant, discoloration could be kept on the brown to black tubers (Figure 7a), with the bruise spreading systematically to the base of the stem, the middle part of the stem, the leaf midrib, the banana bunches, and finally extending to the young fruits. Splitting a young banana fruit revealed hard and stiff flesh with black discoloration along the edges and in the center of the fruit. In contrast, whether singular or combined, treatments involving secondary metabolites did not exhibit fusarium wilt disease symptoms. The fungal isolates had a 100% identity with the Trichoderma asperellum; meanwhile, bacterial isolates had a 98.46% identity with Pseudomonas aeruginosa. The secondary metabolites of T. asperellum and Pseudomonas aeruginosa efficiently inhibit Fusarium wilt disease. The mixed secondary metabolites of T. asperellum and P. aeruginosa showed a similar efficacity in inhibiting Fusarium wilt disease. The potential effect of this discovery is for wider use of these two antagonists and their metabolites to control Fusarium wilt disease and other plant diseases such as vegetable diseases, fruit crops diseases, and estate crops diseases. On the other side, technologically, the active ingredients of these metabolites can be mass-produced as synthetic metabolites on an industrial scale so that the product's price is affordable for the farming community. Furthermore, the government can take more appropriate policies to use organic substances from biological agents that are not harmful to the environment and, at the same time, to reduce the use of chemical pesticides in controlling plant diseases. Even though the research shows satisfactory results, we found the weaknesses of these metabolites. Firstly, it takes a long time to produce these metabolites, and secondly, their effectiveness may decrease over time because natural degradation of these metabolites may occur. Further, it is undoubtedly complex to find them because they are unavailable in the market and can be more expensive than chemical pesticides. Finally, they can take a more extended response to become effective (slow process) in the field than chemical pesticides, and another question is for how long time the metabolites can be stored.
The fungal isolates had a 100% identity with Trichoderma asperellum; meanwhile, bacterial isolates had a 98.46% identity with Pseudomonas aeruginosa. The single secondary metabolites of Trichoderma asperellum, and Pseudomonas aeruginosa efficiently inhibit Fusarium wilt disease 12 months of research period. The mixed secondary metabolites of Trichoderma asperellum and Pseudomonas aeruginosa showed a similar efficacity in inhibiting Fusarium wilt disease. The results of this research provide new hope for using Trichoderma and Pseudomonas metabolites to control various airborne and soil-borne diseases. Also, they can be tried in various climates, mainly to control white root disease in coffee, nutmeg and clove plants in Indonesia. However, it is also necessary to analyze the cost of control using secondary metabolites. In the practical application of these metabolites is necessary to try in the field through root infusion method in controlling soil-borne pathogens or through using stem injection/infuse for airborne pathogens attack the stems, leaves, and fruit, or could be applied together with liquid fertilizer through the leaves.
The discovery of antagonistic isolates and their metabolites useful in agriculture to control various diseases. In the future, it is a need to study how to increase its efficacy through application synergism between antagonist metabolites and plant elicitors that induce plant defense responses that could increase plant's resistance to the pathogen attack. There are still many other questions that need to be explored regarding the use of metabolites, whether these metabolites are selective for the target pathogens or have a broad spectrum, and if so, they could inhibit beneficial microorganisms in agriculture. Regarding safety in the field, testing whether these metabolites have any side effects on the environment and food is necessary. Studying the long-term stability of metabolite molecules in various soil types and soil moisture is also necessary. Finally, it must be designed to make available metabolite product formulations easy to buy in the market and affordable for farmers.
We thank the Universitas Teuku Umar for contributing funds (Leader: Alfizar) (Grant No.: 003/UN59.7/PT.01.03/2020). The University farm of UTU team helped organize the land used and banana seedlings.
[1] Zou, F., Chunming, T., Zhang, B., Wu, W., Shang, N. (2022). The valorization of banana by-products: nutritional composition, bioactivities, applications, and future development. Foods, 11(20): 3170. https://doi.org/10.3390/foods11203170
[2] Acevedo, S.A., Carrillo, A.J.D., Florez-Lopez, E., Grande-Tovar, C.D. (2021). Recovery of banana waste-Loss from production and processing: A contribution to a circular economy. Molecules, 26(17): 5282. https://doi.org/10.3390/molecules26175282
[3] Hikmah, H., Fadli, F., Fitri, I., Kusuma, H., Maulana, O., Bagio, B. (2023). Determinants of consumer demand and preference on banana chips in Central Aceh District. IOP Conference Series: Earth and Environmental Science, 1183: 012048. https://doi.org/10.1088/1755-1315/1183/1/012048
[4] Kurniawan, M.U., Cahyono, A.E., Sukidin., Kantun., S. (2019). Optimization of superior banana product diversification: Empowerment of poor communities. IOP Conference Series: Earth and Environmental Science, 243: 012089. https://doi.org/10.1088/1755-1315/243/1/012089.
[5] Indonesia, A.R. (2016). Adding value to prime commodities of AgroYIndustry in North. Procedia-Social and Behavioral Sciences, 219: 367-373. https://doi.org/10.1016/j.sbspro.2016.05.057
[6] Maryani, N., Lombard, L., Poerba. Y.S., Subandiyah, S., Crous, P.W., Kema, G.J.H. (2019). Phylogeny and genetic diversity of the banana fusarium wilt pathogen fusarium oxysporum f. sp. cubense in the Indonesian centre of origin. Studies Mycology, 92: 155-194. https://doi.org/10.1016/j.simyco.2018.06.003
[7] Effendi, Y., Pambudi, A., Pancoro, A. (2019). Metagenomic analysis of Fusarium oxysporum f. sp. Cubense-infected soil in banana plantation, Sukabumi, Indonesia. Biodiversitas, 20(7): 1939-1945. https://doi.org/10.13057/biodiv/d200721
[8] Maymon, M., Sela, N., Shpaz, U., Galpaz, N., Freeman, S. (2020). The origin and current situation of Fusarium oxysporum f. sp. cubense tropical race 4 in Israel and the Middle East. Scientific Reports, 10(1): 1590. https://doi.org/10.1038/s41598-020-58378-9
[9] Pegg, K.G., Coates, L.M., O'Neill, W.T., Turner, D.W. (2019). The epidemiology of Fusarium Wilt of Banana. Frontier in Plant Science, 10: 1395. https://doi.org/10.3389/fpls.2019.01395
[10] Cannon S, William K., Sreedhar, K., Martin, S., Sarah, J.G., Gero, S. (2022). Multi-site fungicides suppress banana Panama disease, caused by Fusarium Oxysporum f. sp. cubense tropical race 4. PLoS Pathogens, 18(10): e1010860. https://doi.org/10.1371/journal.ppat.1010860
[11] Jamil, A., Shabbir, A. (2020). Utilization of chemical fungicides in managing the wilt disease of chickpea caused by fusarium oxysporum f. sp. ciceri. Archives of Phytopathology and Plant Protection, 53(17-18): 876-898. https://doi.org/10.1080/03235408.2020.1803705
[12] Ismail, A., Khairulmazmi, A., Yasmeen, S., Muhammad, A.A.W., Abdulaziz, B.K., Adamu, A., Syazwan, A.M., Arifin, A., Siti, N.A.A. (2023). Fusarium wilt of banana: current update and sustainable disease control using classical and essential oils approaches. Horticultural Plant Journal, 9(1): 1-28. https://doi.org/10.1016/j.hpj.2022.02.004
[13] Bubici, G., Kaushal, M., Prigigallo, M.I., Cabanas, C.G., Mercado-Blanco, J. (2019). Biological control agents against fusarium wilt of banana. Front Microbiol, 10: 616. https://doi.org/10.3389/fmicb.2019.00616
[14] Guzmán-Guzmán, P., Kumar, A., Santos-Villalobos S.D.L., Parra-Cota, F.I., Orozco-Mosqueda, M.D.C., Fadiji, A.E., Hyder, S., Babalola, O.O., Santoyo, G. (2023). Trichoderma species: Our best fungal allies in the biocontrol of plant diseases - A Review. Plants, 12: 432. https://doi.org/10.3390/plants12030432
[15] Jamil, A., Musheer, N., Kumar, M. (2021). Evaluation of biocontrol agents for management of wilt disease of tomato incited by Fusarium oxysporum f. sp. lycopersici. Archives of Phytopathology and Plant Protection, 54(19-20): 1722-1737. https://doi.org/10.1080/03235408.2021.1938353
[16] Sudirman, A., Sumardiyono, C., Widyastuti, S.M. (2011). Pengendalian Hayati Penyakit Layu Fusarium Pisang (Fusarium oxysporum f. sp. cubense) dengan Trichoderma sp. Jurnal Perlindungan Tanaman Indonesia, 17(1): 31-35. https://doi.org/10.22146/jpti.9396
[17] Puspita, F., Aqita, D.K., Yuda, P.A. (2023). Effectiveness of bio fungicide with active ingredients consortium of Trichoderma virens to control Ganoderma boninense in palm oil nursery. IOP Conference Series: Earth and Environmental Science, 1188(1): 012005. https://doi.org/10.1088/1755-1315/1188/1/012005
[18] Nurbailis, N., Djamaan, A., Rahma, H., Liswarni, Y. (2019). Potential of culture filtrate from Trichoderma spp. as biofungicide to Colletotrichum gloeosporioides causing anthracnose disease in chili. Biodiversitas. Journal of Biological Diversity, 20(10): 2915-2920. https://doi.org/10.13057/biodiv/d201020
[19] Bayoumi, Y., Taha, N., Shalaby, T., Alshaal, T., El-Ramady, H. (2019). Sulfur promotes biocontrol of purple blotch disease via Trichoderma spp. and enhances the growth, yield and quality of onion. Applied Soil Ecology, 134: 15-24. https://doi.org/10.1016/j.apsoil.2018.10.011
[20] Manzar, N., Kashyap, A.S., Goutam, R.S., Rajawat, M.V.S., Sharma, P.K., Sharma, S.K., Singh, H.V. (2022). Trichoderma: Advent of versatile biocontrol agent, its secrets and insights into mechanism of biocontrol potential. Sustainability, 14: 12786. https://doi.org/10.3390/su141912786
[21] Müller, T., Behrendt, U, (2021). Exploiting the biocontrol potential of plant-associated pseudomonads - a step towards pesticide-free agriculture? Biological Control, 155: 104538. https://doi.org/10.1016/j.biocontrol.2021.104538
[22] Banerjee, S., Mukherjee, B., Dutta, S. (2019) Characterization of plant growth promoting rhizobacterial strain Pseudomonas aeruginosa MCC 3877 with special reference to Indole Acetic Acid (IAA) production. International Journal of Pharm Bio Sciences, 10(3): 80-88. https://doi.org/10.22376/ijpbs.2019.10.3.b80-88
[23] Ln, N., Tao, C., Ou, Y., Wang, J., Deng, X., Liu, H., Shen, Z., Li, R., Shen, Q. (2023). Root-associated antagonistic pseudomonas spp. contribute to soil suppressiveness against banana Fusarium wilt disease of banana. Microbiology Spectrum, 11(2): e03525-22. https://doi.org/10.1128/spectrum.03525-22
[24] Russo, C.A.D.M., Selvatti, A.P. (2018). Bootstrap and rogue identification tests for phylogenetic analyses. Molecular Biology and Evolution, 35(9): 2327-2333. https://doi.org/10.1093/molbev/msy118
[25] Stackebrandt, E., Goebel, B.M. (1994). Taxonomic note: A place for DNA-DNA reassociation and 16S rRNA sequence analysis in the present species definition in bacteriology. International Journal of Systematic and Evolutionary Microbiology, 44(4): 846-849. https://doi.org/10.1099/00207713-44-4-846
[26] Rahman, S.S.M.S.A., Zainuddin, N.A.I.M., Aziz, N.A.A. (2021). Evaluation of Trichoderma Asperellum B1902 in controlling fusarium wilt of Cavendish Banana cultivar. Sains Malaysiana, 50(9): 2549-2561. https://doi.org/10.17576/jsm-2021-5009-05
[27] Antari, N.M., Darmayasa, I.B.G., Hardini, J. (2020). Effectiveness of Trichoderma asperellum TKD with mediator of manure to control fusarium disease on red pepper plant (Capsicum annum L.). Simbiosis, 8(2): 63-71.
[28] Soesanto, L., Mugiastuti, E., Rahayuniati, R.F. (2010). Antagonistic mechanisms study of Pseudomonas flourescens P60 on Fusarium oxysporum f.sp. lycopersici of tomato in vivo. Jurnal Hama dan Penyakit Tumbuhan Tropika, 10(2): 108-115.
[29] Aprilia, I., Awang, M., Sobir, S., Suryo, W. (2020). Genetic diversity and fusarium wilt disease resistance (Fusarium oxysporum f. sp cepae) of Indonesian shallots (Allium cepa L. var aggregatum). Jurnal Hortikutura Indonesia, 11(1): 32-40. https://doi.org/10.29244/jhi.11.1.32-40
[30] Manzar, N., Kashyap, A.S., Goutam, R.S., Rajawat, M.V.S., Sharma, P.K., Sharma, S.K., Singh, H.V. (2022). Trichoderma: Advent of versatile biocontrol agent, its secrets and insights into mechanism of biocontrol potential. Sustainability, 14(19): 12786. https://doi.org/10.3390/su141912786
[31] Dukare, A., Paul, S., Asha, S. (2020). Isolation and efficacy of native chitinolytic rhizobacteria for biocontrol activities against fusarium wilt and plant growth promotion in Pigeon pea (Cajanus cajan L.). Egyptian Journal of Biological Pest Control, 30: 56. https://doi.org/10.1186/s41938-020-00256-7
[32] Adriansyah, A., Arri, M.S., Hamawi, M., Ikhwan, A. (2015). Secondary metabolite assay as in vitro antimicrobial of Pseudomonas solanacearum. Gontor Agrotech Science Journal, 2(1): 19-30. https://doi.org/10.21111/agrotech.v2i1.294
[33] Harni, R., Amaria, W., Mahsunah, A.H., Lakanai, I. (2019). Effect of Trichoderma spp. secondary metabolites and botanical fungicide to control VSD disease in cacao. Jurnal Tanaman Industri Penyegar, 6(3): 109-118. https://doi.org/10.21082/jtidp.v6n3.2019.p109-118