Vita Meylani*![]() | Diana Hernawati
| Diana Hernawati![]() | Rinaldi Rizaldi Putra
| Rinaldi Rizaldi Putra![]() | Andri Wibowo
| Andri Wibowo![]()
© 2023 IIETA. This article is published by IIETA and is licensed under the CC BY 4.0 license (http://creativecommons.org/licenses/by/4.0/).
OPEN ACCESS
Coral ecosystems are shaped by complex environmental gradients, and understanding these influences is critical for conservation efforts. This study presents a quantitative analysis of environmental factors contributing to coral species richness along Pangandaran Beach, situated on the southern coast of Java Island within the Indian Ocean—a region noted for its warm, clear waters conducive to coral proliferation. Utilizing line intercept transect surveys for coral assessment and Principal Component Analysis (PCA) for data interpretation, this research identifies a marked variation in species richness between the western (Pasir Putih) and eastern (Batu Numpang) sectors of the beach. The Pasir Putih area exhibits a robust positive correlation between coral species richness and environmental parameters such as dissolved oxygen (DO), light intensity, and water temperature. Conversely, Batu Numpang is characterized by lower species richness, which the Akaike information criterion model (AICc = -236.55) suggests is adversely affected by reduced DO levels—a stark contrast to the positive influence of DO in Pasir Putih (AICc = -14.06). These findings position DO as a pivotal environmental factor influencing coral diversity in Pangandaran Beach, with implications for targeted marine conservation strategies.
Akaike, coral, DO, environmental gradient, PCA
Elucidating the factors driving organismal variation across environmental gradients remains a cornerstone of ecological inquiry [1]. Coral reefs, invaluable yet vulnerable marine ecosystems, have suffered extensively from anthropogenic stresses including overfishing, pollution, disease, and climate-induced bleaching events [1-4]. The Indo-West Pacific and western Atlantic tropical coastal seas have witnessed a pronounced decline in coral cover over the past four decades, with some reefs experiencing shifts to macroalgae-dominated states, commonly referred to as "coral-algal shifts" [4-6]. The depletion of herbivorous fish has been implicated in the top-down degradation of reefs, while bottom-up degradation has been associated with increased nutrient enrichment [7, 8]. Moreover, selective mortality induced by mass bleaching events has contributed to the altered composition of coral assemblages, exacerbating the decline of certain species [9].
Reef composition is influenced by pronounced gradients of depth, salinity, nutrient concentration, and sedimentation from inshore to offshore locations [4, 10-12]. Chronic anthropogenic stressors, such as fishing activities and the discharge of heavy metals and other contaminants, alongside episodic natural disturbances like storms, have been documented to have significant impacts on coral reef populations, with many stressors exhibiting distinct spatial patterns [13]. Interactions among coral taxa further complicate these patterns, influencing community composition and the dynamics of associated taxa [14-16].
Understanding spatial variability and environmental dependencies of coral reef communities is essential for assessing management strategies and sustainable exploitation of natural products [17]. This study aims to examine coral reef taxonomic composition along a significant inshore-offshore gradient at Pangandaran Beach. Data were collected on coral assemblages, and environmental parameters such as water clarity, pH, and dissolved oxygen concentration were measured. The intention is to model the relationship between coral community composition and environmental gradients, providing a robust framework for the prioritization and management of key environmental factors for coral conservation.
2.1 Study location
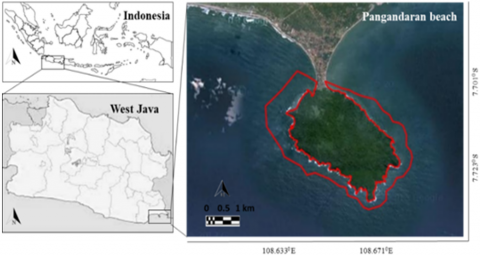
Pangandaran Beach is a beach facing the Indian Ocean. The 530-hectare protected forest known as the Pananjung Nature Reserve is located on an island that extends into the ocean. Pangandaran Beach is separated between western and eastern sections that encircle Pananjung (Figure 1). This study involves sampling in the western portions of the Pasir Putih (PP) area and the eastern portions of the Batu Numpang (BN) area within the longitude geocoordinates of 108.6330 E and 108.6710 E and latitude of 7.7010 S and 7.7230 S.
Figure 1. Map of the study area at Pangandaran beach, West Java Province, Indonesia
2.2 Environmental data collections
Primarily, environmental data were including light (lux), humidity (%), pH, salinity (‰), air temperature (℃), dissolved oxygen (DO, mg/l), wind (m/s), and water temperature (℃) obtained from the western portions of the Pasir Putih area and the eastern portions of the Batu Numpang area within Pangandaran beach. This required employing the line intercept transect survey technique to determine the coral colony coverage [18].
2.3 Coral surveys
During line intercept transect surveys, ocular identifications of scleractinian coral species were made [18]. A method by study [19] was accustomed to photographing unidentified specimens for further analysis and classification. Using the line-intercept transect data, the percentage coverage of each coral species and life form was determined. We surveyed six 30-meter-long transects and placed a 1 m × 1 m quadrat with 1 m spacing at each of the two depths assessed at each site (3 and 5 m). Four locations were studied for corals.
2.4 Coral diversity analysis
The diversity of coral species was determined using The Shannon–Wiener diversity index (H’). This commonly used index was applied to estimate fish species diversity in both disturbed and intact sites.
$H^{\prime}=\sum_{i=1}^S p i \ln (p i)$
where, H’ is Shannon–wiener index, pi is the proportion of the total abundance of the community represented by the i-th species, ln(pi) is the natural log of pi, S = number of fish species encountered, and Σ = sum of species 1 to species S.
2.5 PCA analysis, ANOVA, and x2 test
Principal component analysis (PCA), an ordination method, was used to visualize the patterns of coral species and environmental variable data between western portions of the Pasir Putih area and the eastern portions of the Batu Numpang area. To test the significance of the locations (western vs eastern) on the coral species and environmental variable data, analysis of variance (ANOVA) and x2 test was used.
2.6 AIC model
Coral community correlations with environmental variable data in both western portions of the Pasir Putih area and the eastern portions of the Batu Numpang area were modeled using the Akaike Information Criterion (AIC). The AIC was developed using linear regression. The measured parameters included in AIC were AICc, ΔAICc, AICc weight, cum weight, and log likelihood.
AIC = 2k – 2ln(Ĺ)
with k be the number of estimated parameters in the model. Let Ĺ be the maximized value of the likelihood function for the model.
To build the model, environmental variables correlating with the fish coral community, including light, humidity, pH, salinity, air temperature, dissolved oxygen (DO), wind, and water temperature obtained were included in the analysis to develop the model. The best model was selected based on the model that has the lowest AIC values. To investigate which is the best model, the following three linear mixed-effect models were applied and compared as follows:
Model 1: Light effect
Model 2: Humidity effect
Model 3: pH effect
Model 4: Salinity effect
Model 5: Air temperature effect
Model 6: DO effect
Model 7: Wind effect
Model 8: Water temperature effect
The best model was selected based on the lowest AICc and cum weight values.
Western area (Pasir Putih) of Pangandaran has more coral families and individuals in comparison to eastern area (Batu Numpang). The diversity index for Pasir Putih was 1.494 and 1.561 for Batu Numpang (Table 1). Pocilloporidae was the common coral family since it can be found in both areas. Acroporidae dominated the Batu Numpang, while Pocilloporidae dominated the Pasir Putih. Batu Numpang has more species categorized as near threatened and vulnerable indicating the Batu Numpang was potential for coral conservation areas.
3.1 Environmental gradients
Both sides either the western (Pasir Putih/PP area) and the eastern (Batu Numpang/BN area) have distinct environmental gradients (Table 2). The results of temperature measurements in the waters at the Batu Numpang area showed that water temperature at 27.5℃. Meanwhile, the temperature in Pasir Putih is between 28℃ and 30℃ or warmer than Batu Numpang (Figure 2), with the average temperature of the two points at 28.65℃. According to Keputusan Menteri Lingkungan Hidup dan Kehutanan RI [20], the criteria for appropriate temperature conditions for coral reef life is within the range of 28-30℃. This temperature condition can be classified as normal. The temperature of Batu Numpang and Pasir Putih surpasses the ideal limit for coral growth on average, although corals can still endure it, resulting in a low level of coral cover. Coral reefs thrive in water temperatures ranging from 18 to 36℃, with 26 to 28℃ being the ideal temperature for coral growth. Diverse distribution patterns and coral reef variety are linked to various temperatures in each place, both ecologically and geographically [20, 21].
The salinity value in the Batu Numpang area was 3.23‰, meanwhile, at the Pasir Putih area, the salinity was lower at 3.02‰ (Figure 2). This indicates the salinity levels recorded in both locations were actually not ideal, considering that the recommended salinity levels for coral growth were in the range of 33-34‰ [20].
Table 1. Coral species list recorded with family, common name, abundance, and IUCN Red List in the western (Pasir Putih/PP area) and the eastern (Batu Numpang/BN area)
|
Scientific Name |
Common Name |
Photo |
Family |
IUCN |
Sites |
|
|
Western (PP) |
Eastern (BN) |
|||||
|
Acropora muricata |
staghorn coral |
Acroporidae |
NT |
- |
2 |
|
|
Acropora abrotanoides |
|
LC |
- |
1 |
||
|
Acropora kirstyae |
|
V |
- |
1 |
||
|
Fungia fungites |
common mushroom coral |
Fungiidae |
NT |
- |
1 |
|
|
Pocillopora damicornis |
cauliflower coral |
Pocilloporidae |
LC |
3 |
1 |
|
|
Pocillopora verrucose |
|
|
2 |
- |
||
|
Porites attenuata |
stony coral |
Poritidae |
V |
1 |
- |
|
|
Pavona frondifera |
leaf coral |
Agariciidae |
LC |
1 |
- |
|
|
Dipsastraea pallida |
colonial stony coral |
Merulinidae |
LC |
1 |
- |
|
|
Number of coral families |
4 |
3 |
||||
|
Number of coral species |
5 |
5 |
||||
|
Number of coral individuals |
8 |
6 |
||||
|
Shannon diversity index (H’) |
1.494 |
1.561 |
||||
IUCN Red List status: LC (least concern), NT (near threatened), V (vulnerable)
Table 2. Summary of the mean and 95%CI and ANOVA for the effects of locations (western vs eastern of Pangandaran beach) on the coral species, light (lux), humidity (%), pH, salinity (‰), air temperature (℃), dissolved oxygen (DO, mg/l), wind (m/s), and water temperature (℃)
|
Source of Variations |
Mean and 95%CI |
F |
P |
|
|
Western (PP) |
Eastern (BN) |
|||
|
Species |
4(95%CI: 1.74 to 6.26) |
3(95%CI: 1.87 to 4.13) |
0.6 |
0.481817415 |
|
Light (lux) |
815.4(95%CI: 779 to 851) |
769(95%CI: 487 to 1050) |
0.102473628 |
0.764905066 |
|
Humidity (%) |
83(95%CI: 80 to 86) |
85(95%CI: 85 to 85) |
1.714285714 |
0.260574547 |
|
pH |
6.4(95%CI: 6.17 to 6.63) |
6.3(95%CI: 6.07 to 6.53) |
0.375 |
0.573392254 |
|
Salinity (‰) |
3.146(95%CI: 3.02 to 3.28) |
3.23(95%CI:3.23 to 3.23) |
1.714285714 |
0.260574547 |
|
Air Temperature (℃) |
30.4(95%CI: 30.2 to 30.6) |
30.3(95%CI: 30.1 to 30.5) |
0.375 |
0.573392254 |
|
Dissolved Oxygen (DO, mg/l) |
5.48(95%CI: 4.65 to 6.31) |
5(95%CI: 4.21 to 5.79 |
0.675527756 |
0.45729822 |
|
Wind (m/s) |
4.278(95%CI: 2.96 to 5.6) |
5.67(95%CI: 4.78 to 6.56 |
2.924905354 |
0.162395957 |
|
Water temperature (℃) |
28.42(95%CI: 27.5 to 29.3) |
27.5(95%CI: 27.5 to 27.5) |
4.368891948 |
0.104808273 |
Figure 2. Comparisons of coral species, light (lux), humidity (%), pH, salinity (‰), air temperature (℃), dissolved oxygen (DO, mg/l), wind (m/s), and water temperature (℃) between the western (Pasir Putih/PP area) and the eastern (Batu Numpang/BN area)
The weather and the time of sampling might alter the salinity levels, which can affect the salinity of the waters. For corals, salinity is recognized to be a limiting factor [21]. According to Supriharyono [22], the typical salinity of saltwater in the tropics is around 35‰, and corals grow in the 34-36‰ salinity range. Salinity, on the other hand, is a significant element in aquatic ecological circumstances that can influence osmotic pressure in the aquatic organism's body, causing these creatures to waste energy to adapt to their surroundings via osmoregulation systems [23, 24].
Gradients in water quality were more important predictors of variance in coral composition. As can be seen in PCA model (Figure 3), the biotic composition of selected coral taxa (ascidians, macroalgae, and corals) on the Pangandaran beach seems to be influenced by water quality characteristics include light intensity (turbidity), salinity, water temperature and DO, which is consistent with Gittenberger et al. [25]. According to the Akaike model (Table 3), declining DO is a main determinant factor (AICc = -14.06) contribute to the increases of coral species in Pasir Putih and decline in Batu Numpang (AICc = -236.55). To conclude, DO gradient is key environmental factors influencing the coral community in the Pangandaran beach. This is in contrast to Cleary et al. [10], who investigated coral, sponge, large benthic foraminifera (LBF), and sediment archaea and bacteria populations in the Spermonde archipelago. With the exception of soil bacteria, habitat variables were more relevant explanatory factors than water quality variables in the Spermonde. The current study, in which water quality variables were more important predictors of ascidians' composition than habitat variables, is consistent with Gittenberger et al. [25], who found that various ascidian species were very good indicators for bioregions with varying distances offshore, while no correlations between ascidian communities and habitats were found.
Larger foraminifera harboring symbionts are among the most essential calcifiers in healthy reef environments [26, 27]. They are sensitive to environmental factors and are commonly employed as water quality monitors [28, 29]. As seen in inshore reefs, poor water quality results in fewer obligate symbiont-bearing taxa compared to non-symbiont-bearing taxa [28]. Depth is a key factor in defining the assemblage composition of LBF within reef systems, with the biggest diversity in assemblage composition occurring in either shallow (reef flat) or deeper (reef slope) assemblages [30, 31].
Figure 3. PCA models showing the western (Pasir Putih/PP area) and the eastern (Batu Numpang/BN area) related to light, humidity (%), pH, salinity, air temperature (AT), dissolved oxygen (DO), wind velocity (WV), and water temperature (WT)
The abundance of Amphistegina species determines a major part of the variation in the fraction of symbiont-bearing LBF in the total benthic LBF assemblage. However, there are more LBF species in the Indo-West Pacific, and they occupy a wider range of habitats [30-32]. Certain taxa in the LBF, particularly those in the genus Calcarina, can endure both poor water quality and benthic environments dominated by algae rather than corals [31]. Some species in this group are particularly sensitive to substrate types, such as the presence of corals and the lack of sand, while others are sensitive to water quality [31]. Reefs at JBTI are dominated by Calcarina species that withstand poor water quality, in contrast to other reef systems in the region. In addition, taxa that tolerate algal rather than rubble substrates predominated. We found that water quality appears to be a key determinant of LBF assemblage composition in this study.
In this study, modeling the coral reef compositions along with their environmental gradients will provide versatile coral management tools. Since it can determine which environmental variables should be prioritized and managed, this can lead to effective coral reef management.
Table 3. AIC models of coral species, light, humidity, pH, salinity, air temperature, dissolved oxygen (DO), wind, and water temperature between the western (Pasir Putih/PP area) and the eastern (Batu Numpang/BN area)
|
Coral Species Richness (S) Models |
AICc |
ΔAICc |
AICc Weight |
Cum. Weight |
Log likelihood |
|
Batu Numpang (East) |
|
|
|
|
|
|
Model 1: S’⁓light* |
-236.55 |
0.00 |
0.92 |
0.92 |
109.27 |
|
Model 2: S’⁓humidity |
-11.57 |
224.98 |
0.00 |
1.00 |
-3.22 |
|
Model 3: S’⁓pH |
-11.57 |
224.98 |
0.00 |
1.00 |
-3.22 |
|
Model 4: S’⁓salinity |
-11.57 |
224.98 |
0.00 |
1.00 |
-3.22 |
|
Model 5: S’⁓air temp |
-11.57 |
224.98 |
0.00 |
1.00 |
-3.22 |
|
Model 6: S’⁓DO* |
-227.08 |
9.46 |
0.01 |
1.00 |
104.54 |
|
Model 7: S’⁓wind velocity* |
-231.36 |
5.16 |
0.07 |
0.99 |
106.68 |
|
Model 8: S’⁓water temp |
-11.57 |
224.98 |
0.00 |
1.00 |
-3.22 |
|
Pasir Putih (West) |
|
|
|
|
|
|
Model 1: S’⁓light |
-9.35 |
4.73 |
0.03 |
0.97 |
-4.32 |
|
Model 2*: S’⁓humidity* |
-13.24 |
0.84 |
0.23 |
0.82 |
-2.38 |
|
Model 3*: S’⁓pH |
-7.41 |
6.68 |
0.01 |
0.99 |
-5.30 |
|
Model 4: S’⁓salinity* |
-13.24 |
0.84 |
0.23 |
0.59 |
-2.38 |
|
Model 5: S’⁓air temp |
-7.41 |
6.68 |
0.01 |
1.00 |
-5.30 |
|
Model 6: S’⁓DO* |
-14.06 |
0.00 |
0.36 |
0.36 |
-1.96 |
|
Model 7: S’⁓wind velocity |
-10.51 |
3.57 |
0.06 |
0.88 |
-3.74 |
|
Model 8: S’⁓water temp |
-10.44 |
3.64 |
0.06 |
0.94 |
-3.76 |
*= best model
Water quality and environmental gradients have shaped the coral community in Pangandaran beach in particular Pasir Putih in western sides and Batu Numpang in eastern sides of beach. The diversity index for Pasir Putih was 1.494 and 1.561 for Batu Numpang, indicating high coral diversity mainly in Batu Numpang. Pocilloporidae was the common coral family since it can be found in both Pasir Putih and Batu Numpang. Acroporidae dominated the Batu Numpang, while Pocilloporidae dominated the Pasir Putih. Batu Numpang has more species categorized as near threatened and vulnerable indicating the Batu Numpang was potential for coral conservation areas., decline in Batu Numpang (AICc = -236.55). To conclude, DO gradient is key environmental factors influencing the coral community in the Pangandaran Beach.
Despite the fact that the study has succeeded in modeling the coral community across environmental gradients, for future studies, it is important to include more environmental factors to fill data gaps. Those factors and data requirements include the nutrient level, sedimentation, and turbidity.
We are grateful to LP2MPMP Universitas Siliwangi for granting research permits. We thank staff of Biology Education Laboratory (Ari Hardian & Asep Yudi Supriatna) for support this research. We also thank B.W. Hoeksema and the staff of Groningen Institute for Evolutionary Life Sciences and Naturalis Biodiversity Center Darwinweg for supporting this research and identifying the coral species.
[1] Mora, C., Chittaro, P.M., Sale, P.F., Kritzer, J.P., Ludsin, S.A. (2003). Patterns and processes in reef fish diversity. Nature, 421: 933-936.
[2] Roberts, C.M., McClean, C.J., Veron, J.E., et al. (2002). Marine biodiversity hotspots and conservation priorities for tropical reefs. Science, 295(5558): 1280-1284. https://doi.org/10.1126/science.1067728
[3] Walther, G.R., Post, E., Convey, P., et al. (2002). Ecological response to recent climate change. Nature, 416: 389-395.
[4] Hoey, A.S., Bellwood, D.R. (2008). Cross-shelf variation in the role of parrotfishes on the Great Barrier Reef. Coral Reefs, 27: 37-47. https://doi.org/10.1007/s00338-007-0287-x
[5] Ansell, A., Gibson, R., Barnes, M. (1998). Ecological impact of green macroalgal blooms. Oceanography and Marine Biology: An Annual Review, 36: 97-125.
[6] Schaffelke, B., Mellors, J., Duke, N.C. (2005). Water quality in the Great Barrier Reef region: Responses of mangrove, seagrass and macroalgal communities. Marine Pollution Bulletin, 51(1-4): 279-296. https://doi.org/10.1016/j.marpolbul.2004.10.025
[7] Bruno, J.F., Precht, W.F., Vroom, P.S., Aronson, R.B. (2014). Coral reef baselines: How much macroalgae is natural? Marine Pollution Bulletin, 80(1-2): 24-29. https://doi.org/10.1016/j.marpolbul.2014.01.010
[8] Burkepile, D.E., Allgeier, J.E., Shantz, A.A., Pritchard, C.E., Lemoine, N.P., Bhatti, L.H., Layman, C.A. (2013). Nutrient supply from fishes facilitates macroalgae and suppresses corals in a Caribbean coral reef ecosystem. Scientific Reports, 3: 1493. https://doi.org/10.1038/srep01493
[9] Carpenter, K.E., Abrar, M., Aeby, G., et al. (2008). One-third of reef-building corals face elevated extinction risk from climate change and local impacts. Science, 321(5888): 560-563. https://doi.org/10.1126/science.1159196
[10] Cleary, D.F., Polónia, A.R., Renema, W., Hoeksema, B.W., Wolstenholme, J., Tuti, Y., de Voogd, N.J. (2014). Coral reefs next to a major conurbation: A study of temporal change (1985-2011) in coral cover and composition in the reefs of Jakarta, Indonesia. Marine Ecology Progress Series, 501: 89-98. https://doi.org/10.3354/meps10678
[11] Cleary, D.F., De Vantier, L., Giyanto, Vail, L., Manto, P., de Voogd, N.J., Rachello-Dolmen, P.G., Tuti, Y., Budiyanto, A., Wolstenholme, J., Hoeksema, B.W. (2008). Relating variation in species composition to environmental variables: A multi-taxon study in an Indonesian coral reef complex. Aquatic Sciences, 70: 419-431. https://doi.org/10.1007/s00027-008-8077-2
[12] Cleary, D.F.R., Suharsono, Hoeksema, B.W. (2006). Coral diversity across a disturbance gradient in the Pulau Seribu reef complex off Jakarta, Indonesia. In: Marine, Freshwater, and Wetlands Biodiversity Conservation, Springer, Dordrecht, pp. 285-306. https://doi.org/10.1007/978-1-4020-5734-2_19
[13] Goatley, C.H.R., Bellwood, D.R. (2013). Ecological consequences of sediment on high-energy coral reefs. PLOS ONE, 8(10): e77737. https://doi.org/10.1371/journal.pone.0077737
[14] Bellwood, D.R., Hughes, T.P., Folke, C., Nystro, M. (2004). Confronting the coral reef crisis D. Nature, 429: 827-833. https://doi.org/10.1007/978-3-540-25937-4_9
[15] Berumen, M.L., Pratchett, M.S. (2006). Recovery without resilience: Persistent disturbance and long-term shifts in the structure of fish and coral communities at Tiahura Reef, Moorea. Coral Reefs, 25: 647-653. https://doi.org/10.1007/s00338-006-0145-2.
[16] Hughes, T.P., Graham, N.A.J., Jackson, J.B.C., Mumby, P.J., Steneck, R.S. (2010). Rising to the challenge of sustaining coral reef resilience. Trends in Ecology & Evolution, 25(11): 633-642. https://doi.org/10.1016/j.tree.2010.07.011
[17] Tuomisto, H., Ruokolainen, K., Yli-Halla, M. (2003). Dispersal, environment, and floristic variation of Western Amazonian forests. Science, 299(5604): 241-244. https://doi.org/10.1126/science.1078037
[18] English, S., Wilkinson, C., Baker, V. (1997). Survey Manual for Tropical Marine Resources, Townsville, Australia: Australian Institute of Marine Science, 1997.
[19] Kurniawan, R., Ariestasari, A., Silalahi, R.S., et al. (2019). Identification Acroporidae and Favidae by a newly approach called Reef Identification Knowhow Application-Reconstructed by 3D Imagery (RIKA-R3DI) Method,” MethodsX, 6: 1084-1100. https://doi.org/10.1016/j.mex.2019.05.002
[20] Keputusan menteri LHK. https://ppkl.menlhk.go.id/website/filebox/824/191009100640Keputusan%20MENLH%20Nomor%2051%20tahun%202004%20%20tentang%20Baku%20Mutu%20Air%20Laut.pdf, accessed on Dec. 20, 2023.
[21] Tingkat tutupan ekosistem terumbu karang di perairan pulau terkulai. https://www.researchgate.net/publication/322055916_TINGKAT_TUTUPAN_EKOSISTEM_TERUMBU_KARANG_DI_PERAIRAN_PULAU_TERKULAI, accessed on Nov. 26, 2023.
[22] Pengelolaan ekosistem terumbu karang. https://onesearch.id/Record/IOS3239.slims-42235?widget=1, accessed on Dec. 20, 2023.
[23] Hallock, P., Lidz, B.H., Cockey-Burkhard, E.M., Donnelly, K.B. (2003). Foraminifera as bioindicators in coral reef assessment and monitoring: the FORAM index. Environmental Monitoring and Assessment, 81: 221-238. https://doi.org/10.1023/A:1021337310386
[24] Legendre, P., Gallagher, E.D. (2001). Ecologically meaningful transformations for ordination of species data. Oecologia, 129: 271-280. https://doi.org/doi: 10.1007/s004420100716
[25] Gittenberger, A., Draisma, S.G., Arbi, U.Y., Langenberg, V., Erftemeijer, P.L., Tuti, Y., Hoeksema, B.W. (2015). Coral reef organisms as bioregion indicators off Halmahera, Moluccas, Indonesia. Aquatic Conservation: Marine and Freshwater Ecosystems, 25(6): 743-755. https://doi.org/10.1002/aqc.2495
[26] Scoffin, T.P., Tudhope, A.W. (1985). Sedimentary environments of the central region of the great barrier reef of Australia. Coral Reefs, 4: 81-93. https://doi.org/10.1080/14702549308521159
[27] Renema, W. (2006). Large benthic foraminifera from the deep photic zone of a mixed siliciclastic-carbonate shelf off East Kalimantan, Indonesia. Marine Micropaleontology, 58(2): 73-82. https://doi.org/10.1016/j.marmicro.2005.10.004
[28] Ayoub, L., Hallock, P., Coble, P. (2008). Colored dissolved organic material increases resiliency of coral reefs by controlling exposure to UVR. In Proceedings of the 11th International Coral Reef Symposium, Ft. Lauderdale, Florida, pp. 572-576.
[29] Uthicke, S., Nobes, K. (2008). Benthic foraminifera as ecological indicators for water quality on the Great Barrier Reef. Estuarine, Coastal and Shelf Science, 78(4): 763-773. https://doi.org/10.1016/j.ecss.2008.02.014
[30] Hohenegger, J. (1994). Distribution of living larger foraminifera NW of Sesoko-Jima, Okinawa, Japan. Marine Ecology, 15(3-4): 291-334. https://doi.org/10.1111/j.1439-0485.1994.tb00059.x
[31] Renema, W. (2010). Is increased calcarinid (foraminifera) abundance indicating a larger role for macro-algae in Indonesian Plio-Pleistocene coral reefs? Coral Reefs, 29: 165-173. https://doi.org/10.1007/s00338-009-0568-7
[32] Renema, W. (2008). Habitat selective factors influencing the distribution of larger benthic foraminiferal assemblages over the Kepulauan Seribu. Marine Micropaleontology, 68(3-4): 286-298. https://doi.org/10.1016/j.marmicro.2008.06.002