Influence of Rhizosphere-Isolated Indigenous Bacteria on Growth and Yield of Soybean (Glycine max L.) Devon 2 Varieties in Mugarsari Land

Dedi Natawijaya*![]() | Ida Hodiyah
| Ida Hodiyah![]() | Visi Tinta Manik
| Visi Tinta Manik![]()
© 2023 IIETA. This article is published by IIETA and is licensed under the CC BY 4.0 license (http://creativecommons.org/licenses/by/4.0/).
OPEN ACCESS
Indigenous bacteria thriving in natural environments can serve as an alternative to biofertilizers in promoting plant growth. This study aimed to isolate and quantify the abundance of bacteria from the rhizospheres of calopo (Calopogonium mucunoides), reeds (Imperata cylindrica), and kirinyuh (Eupatorium odoratum) in Mugarsari land. Furthermore, the effects of applying isolated nitrogen-fixing bacteria, phosphate-solubilizing bacteria, and organic matter-decomposing bacteria on the growth and yield of soybean (Glycine max L.) Devon 2 varieties were investigated. A randomized block design with five treatments and five replications was employed. Results revealed a diverse range and abundance of bacteria isolated from calopo plants, reeds, and kirinyuh in the Mugarsari land rhizosphere. Bacterial inoculation significantly influenced the number of leaves, total chlorophyll content, the total number of effective root nodules, and the shoot/root ratio in soybean plants. However, plant height, leaf area, root length, the total number of ineffective root nodules, wet weight of plant biomass, number of pods per plant, number of seeds per plant, weight of seeds per plant, and weight of 100 dried seeds were not significantly affected. This study highlights the potential of indigenous bacteria as an eco-friendly alternative in enhancing soybean growth and yield in Mugarsari land.
indigenous, biofertilizer, nitrogen-fixing, phosphate-solubilizing, organic decomposing, soybean
Agriculture plays a vital role in Indonesia's economy, with approximately 30% of the nation's land area allocated for this sector according to the Indonesian Ministry of Agriculture. Over the past few decades, the proportion of agricultural land has increased to 32%, making Indonesia a global leader in producing various tropical agricultural products due to its abundant fertile soils. However, Indonesian farmers and agricultural laborers face challenges in accessing sufficient agricultural territory. Java and Bali, which constitute only 7% of Indonesia's total land area, accommodate 60% of the country's population, resulting in highly intensive agriculture with up to three crop cycles per year [1].
The extensive use of chemical pesticides and fertilizers in Indonesian agriculture has raised concerns regarding their environmental impact and effects on soil quality. Studies have reported that the current application of pesticides and chemical fertilizers affects rice cultivation in rural Java [2]. Despite these concerns, farmers show reluctance towards adopting organic farming techniques. Alta et al. [3] highlighted that the overuse of urea and the underuse of organic fertilizer negatively affect the environment, particularly through soil deterioration (chemical, physical, and biological soil conditions).
Soil ecosystems consist of five principal components: mineral matter, water, organic matter, air, and living organisms. Microorganisms constitute the majority of soil's living organisms and play a crucial role in nutrient production for plants. These microorganisms contribute to the decomposition of organic materials, nutrient cycling, and soil structure improvement [4].
The application of soil microorganisms in agriculture is an environmentally sustainable approach that is technically feasible, commercially advantageous, and socially acceptable [5]. Biofertilizers, also known as biological fertilizers, are soil microbes that support plant growth and productivity. These substances contain beneficial microorganisms that enhance crop yields and soil fertility through increased biological activity, which may interact with the soil's chemical and physical properties [6]. Nitrogen-fixing microorganisms, phosphate-solubilizing microorganisms, and organic matter decomposers are commonly employed as active components in biological fertilizers [7].
Indigenous bacteria can be utilized as biofertilizers to decompose organic debris, assist in mineralization, and symbiotically or freely coexist with plants to anchor or provide nutrients for plant growth. These naturally occurring microorganisms offer numerous benefits, including their potential use as plant control agents, antibiotic producers, phosphate dissolvers, nitrogen fixers, enzyme producers, and soil remediators [8].
The rhizosphere provides an ideal environment for soil microorganism growth and development, supporting their nutrition. Nitrogen-fixing bacteria (NFB) can enhance the efficiency of nitrogen utilization from the soil, while phosphorus-solubilizing bacteria (PSB) can dissolve fixed phosphorus and convert it to a plant-available form. Organic decomposing bacteria (ODB) decompose residual organic matter, returning it to the soil for plant reuse [9, 10].
Soybeans are Indonesia's third most important food crop; however, the country struggles to meet local consumption demands. In 2021, Indonesia imported 2.49 million tonnes of soybeans worth US$ 1.48 billion or Rp. 21.9 trillion [11]. Several challenges contribute to this situation, including intense competition for land with other commodities, low yield stability, limited success in expanding planting areas, poor seed quality, unfavorable trading conditions, and less intensive cultivation techniques.
Previous research has shown that the application of soil microorganisms, such as rhizobium and biological fertilizers, can enhance soybean growth and yield [12]. Studies by Pedrozo et al. [13] and Cordeiro and Echer [14] demonstrated that using nitrogen-fixing bacteria inoculants increased soybean productivity. Despite numerous studies on the use of microbes for soybean growth and yield improvement, the effects of local bacteria on legume growth and yield are less explored. Most studies focus on specific bacterial genotypes, such as Bradyrhizobium and Bacillus, and their effects on soybean growth and yield [15-17]. Further research is required to investigate the potential of local microorganisms in promoting soybean growth and yield, contributing to sustainable agriculture in Indonesia.
This study aimed to identify and evaluate the potential of indigenous bacterial inoculants as a cost-effective, eco-friendly approach to improving soil quality while reducing the reliance on chemical fertilizers. Local bacterial species (indigenous) exhibit greater adaptability than introduced species, making it essential to assess the diversity and abundance of local bacteria for rapid adaptation to the plants cultivated on the land.
Three types of bacteria were identified through laboratory tests in this study: nitrogen-fixing bacteria (NFB), phosphorus-solubilizing bacteria (PSB), and organic decomposing bacteria (ODB). These bacteria were then tested for their effects on soybean plants in the field.
This study employs both descriptive and experimental methodologies. The descriptive approach was used to isolate and identify soil bacteria from the Mugarsari land's rhizosphere. The experimental study was used to examine the effects of the isolated indigenous bacteria from descriptive research on the growth and yield of the Devon 2 soybean (Glycine max L.) variety. The experiment was carried out using a Randomized Block Design (RBD) with five treatments and five replications. The treatment consists of B0 (no inoculation), B1 (inoculation of nitrogen-fixing bacteria (NFB)), B2 (inoculation of phosphate-solubilizing bacteria (PSB), B3 (inoculation of organic decomposing bacteria (ODB), and B4 (inoculation of a mixture of NFB+PSB+ODB).
2.1 Soil sampling
Soil samples were obtained from the Mugarsari campus area of Universitas Siliwangi, Tasikmalaya, West Java, Indonesia. Three samples of soil were collected from three separate sites: the rhizospheres of the Kalopo plant (Calopogonium mucunoides) (Lat: 07°22’50.9106” Long: 108°15’1.4976” Elevation: 367 masl); the Imperata cylindrica plant (Lat: 07°22’51.7066” Long: 108°15’2.5985” Elevation: 367 masl); and the Kirinyuh plant (Eupatorium odoratum) (Lat: 07°22’51.9856” Long: 108°15’3.488” Elevation: 367 masl). Then, soil samples were combined for further bacterial isolation in the lab.
2.2 Bacterial isolation
Indigenous bacteria were isolated by vortexing for two minutes 10 grams of rhizosphere soil composite samples in a sterilized 90 mL 0.85% NaCl solution. After separating the suspension, 1 mL was mixed with 9 mL of 0.85% NaCl solution to make a series of dilutions ranging from 10-1-10-7. Then, 0.1 mL or 100 μL of each dilution series was spread on YEMA-CR selective media for NFB, Pycovskaya Agar for PSB, and Carboxy Methyl Cellulose Congo Red (CMC-CR) media for ODB. Each planting was done twice and incubated for 48 hours at a temperature of 27-28° Celsius. The morphological characteristics of the bacterial colonies were observed using Bergey's manual book, and gram staining was used according to Cappucinno and Sherman [18]. The colonies were counted using the Total Plate Count (TPC) method.
2.3 Bacterial qualitative assay
A qualitative assay was carried out by inoculating bacterial isolate on Pycovskaya Agar for phosphate solubilizing, Carboxy Methyl Cellulose Media for cellulosic activity assay, and Skim Milk (SM) Agar medium for potential proteolytic activity.
2.4 Hypersensitivity assay
A hypersensitivity assay was carried out on tobacco plants. Each bacterial inoculum was injected into the tobacco leaf at a density of 106 CFU/mL using a needless syringe. The outcome was then observed after 2 x 24 hours. If the bacteria exhibit necrosis symptoms, they are pathogenic to plants [19]. Bacteria with the highest ability in nitrogen-fixing activity, phosphate solubilization, and organic decomposition were then applied to soybean plants.
2.5 Bacterial inoculant application
Bacterial inoculant at 106 CFU/mL density was applied by soaking soybean seeds for 1 hour prior to planting and then pouring 10 mL of inoculant on the planting medium at 14 days after planting (dap) and 35 dap [20]. The vegetative growth response of soybeans was observed 14 dap, and the yield was observed 70 days later.
3.1 Bacterial screening
Based on observations of the morphological colony (shape, margin, elevation color, surface, and size) from 3 soil composites, three groups of indigenous bacterial isolates were discovered, i.e., 18 isolates of Nitrogen Fixing Bacteria (NFB), 16 isolates Phosphate Solubilizing Bacteria (PSB) and 36 isolates Organic Decomposing Bacteria (ODB). The majority of the NFB isolates (10 isolates) had a round or circular shape, while 7 isolates had irregular shapes, and 1 isolate had a rhizoid shape. Based on the margin characteristics, there were 10 entire isolates, 7 undulate isolates, and 1 lobate isolate. There were eight pink isolates and ten white isolates. There were seven convex elevation isolates, seven raised isolates, four umbonated isolates, and one flat isolate. The majority (12 isolates) had a rather shiny surface, and 6 isolates had a shiny surface, and comes in three sizes: large (10 isolates), medium (2 isolates), and small (6 isolates).
The circular shapes dominated the PSB isolates (14 isolates), and two isolates were amoeboid in shape. 11 isolates had entire edges, 3 isolates had irregular edges, and 2 isolates had rhizoid edges based on margins. Based on color, there were 8 yellow isolates, 4 cream isolates, and 4 white isolates. The majority of the phosphate-solubilizing bacterial isolates (9 isolates) had a convex elevation, while the remaining 7 isolates had a flat elevation. There are 9 rather shiny isolates and 7 shiny isolates on the surface. There were three large isolates, seven medium-sized isolates, and six small-sized isolates.
ODB isolates are classified into four groups based on their dominant shape: circular (17 isolates), punctiform (15 isolates), irregular (3 isolates), and spindle (1 isolate). Based on the margins, most (33 isolates) had an entire margin, and 3 isolates had a margin of 3 isolates. 29 isolates were cream-colored, and 7 isolates were white. Based on elevation, 25 isolates had convex elevation, 9 isolates had flat elevation, and 2 isolates had non-convex elevation. Based on the surface, there were 18 isolates that had a rather shiny surface, and 18 isolates had a shiny surface.
According to the results of bacterial isolation, indigenous bacterial isolates had a variety of shapes, edges (margins), colors, elevations, surfaces, and sizes (Figure 1). Bacterial colonies have specific properties on solid media [21]. Bacteria have cultural characteristics that classify microorganisms based on group taxonomy based on these differences [18]. Differences in colony morphology suggest that indigenous bacterial colonies are from different species [22]. The genetic composition of organisms determines growth's color, size, shape, and texture; environmental factors also influence nutrient provision, temperature, and incubation time [23].
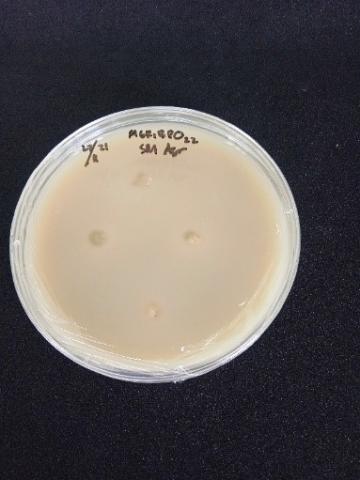
The qualitative screening results revealed that the total isolates were successfully screened based on their highest ability to solubilize phosphate, cellulolytic activity, and proteolytic activity (Figure 2). There were 17 potential bacterial isolates, including four nitrogen-fixing bacteria, eight phosphate-solubilizing bacteria, and five organic degrading bacteria. The isolates were then tested for gram and hypersensitivity. Nitrogen-fixing bacteria have a positive bacilli cell shape, phosphate-solubilizing bacteria have a negative coccus cell shape, and organic decomposing bacteria have a positive-negative coccus shape, according to the gram test.
(a)
(b)
(c)
Figure 1. Indigenous bacterial morphology from Mugarsari land rhizosphere; (a) PSB; (b) NFB and (c) ODP
(a)
(b)
(c)
Figure 2. Qualitative screening of indigenous bacterial isolates; (a) solubilization of phosphate in vitro by indigenous bacterial isolates, the formation of halo zone around the colonies shows the solubilization of inorganic phosphate; (b) cellulose hydrolysis using bacterial isolates, the activity was shown by a clear zone around the colony, and (c) qualitative proteolytic assay was conducted using skim milk as a substrate, the capacity to produce extracellular proteases was indicated by the formation of clear zones around the colony
(a)
(b)
Figure 3. Hypersensitivity test and response to symptoms of necrosis on tobacco leaves; (a) necrosis negative, (b) necrosis positive
Table 1. Hypersensitivity assay of indigenous rhizosphere bacteria isolates from Mugarsari land on tobacco leaves
|
No |
Code of Isolates |
Necrosis |
|
1 |
MGR-BPN-11 |
Negatif |
|
2 |
MGR-BPN-14 |
Negatif |
|
3 |
MGR-BPN-16 |
Negatif |
|
4 |
MGR-BPN-2 |
Negatif |
|
5 |
MGR-BPF-1 |
Negatif |
|
6 |
MGR-BPF-2 |
Positif |
|
7 |
MGR-BPF-4 |
Negatif |
|
8 |
MGR-BPF-5 |
Negatif |
|
9 |
MGR-BPF-6 |
Positif |
|
10 |
MGR-BPF-9 |
Negatif |
|
11 |
MGR-BPF-11 |
Negatif |
|
12 |
MGR-BPF-14 |
Negatif |
|
13 |
MGR-BPO-11 |
Negatif |
|
14 |
MGR-BPO-12 |
Negatif |
|
15 |
MGR-BPO-13 |
Negatif |
|
16 |
MGR-BPO-16 |
Negatif |
|
17 |
MGR-BPO-22 |
Negatif |
Table 2. Effect of indigenous bacterial on soybean plant height
|
Treatment |
Plant height (cm) |
|||
|
14 dap |
21 dap |
28 dap |
35 dap |
|
|
Whithout inoculation |
29.23 a |
41.47 a |
55.25 a |
87.72 a |
|
NFB inoculation |
28.36 a |
40.25 a |
55.90 a |
87.22 a |
|
PSB inoculation |
27.15 a |
40.37 a |
55.80 a |
85.92 a |
|
ODB inoculation |
27.26 a |
41.50 a |
56.42 a |
88.92 a |
|
NFB+PSB+ODB |
27.49 a |
41.14 a |
54.62 a |
85.58 a |
Note: The Duncan's Multiple Range Test at a 5% error level indicates that there is no statistically significant difference between the mean number and the letter that comes after it.
Table 3. Effect of indigenous bacterial on soybean plant number of leaves
|
Treatment |
Number of leaves |
|||
|
14 dap |
21 dap |
28 dap |
35 dap |
|
|
Without inoculation |
3.14 a |
5.08 b |
5.77 a |
11.09 a |
|
NFB inoculation |
3.11 a |
5.04 b |
5.87 a |
11.73 a |
|
PSB inoculation |
3.03 a |
4.93 b |
5.70 a |
10.73 a |
|
ODB inoculation |
2.97 a |
4.28 ab |
5.69 a |
11.47 a |
|
NFB+PSB+ODB |
3.21 a |
4.08 a |
5.84 a |
11.29 a |
Note: The Duncan's Multiple Range Test at a 5% error level indicates that there is no statistically significant difference between the mean number and the letter that comes after it.
The hypersensitivity reaction test on tobacco plant leaves revealed that 15 isolates had a negative reaction (did not cause necrosis symptoms) and 2 isolates had a positive reaction. The hypersensitive response (HR) test determines whether or not a bacterial isolate is pathogenic [24]. The symptoms of tobacco leaf necrosis are depicted in Figure 3.
The hypersensitivity test response is the most crucial test for determining whether bacteria are pathogenic. This test identifies pathogenic gram-positive and gram-negative bacteria [25]. Tobacco plants respond quickly to bacteria tested on leaf tissue; local (necrotic) lesions indicate this response.
According to Table 1, two isolates from the Mugarsari land rhizosphere reacted positively to necrosis, while fifteen isolates reacted negatively. Bacterial isolates that react negatively (do not cause necrosis) are then screened using the qualitative test that has the greatest potential for application to soybeans. According to García-Orenes et al. [26], rapid cell death in and around cells infected with pathogens is a hypersensitive reaction associated with a plant resistance response.
3.2 Soybean vegetative growth
No treatment significantly differed in soybean plant height, presumably due to low nutrient availability, particularly the elements Carbon (C), Nitrogen (N), and Potassium (K), which affect plant growth and microbial activity in the soil (Table 2). Furthermore, competition can cause inoculants to be unable to adapt or associate well with the growing media, resulting in impediments in the functional mechanism of bacteria in spurring plant growth, one of which affects soybean plant growth.
Giving bacteria to the soil as inoculants, according to [27], generally cannot maintain a high population, and their numbers will decrease to a low level. Because indigenous bacteria are adaptable to the plant environment and soil conditions, they compete. Glick [28] believes that competition can reduce bacteria's effectiveness. According to our research, the growth of functional characters of inoculants that promote plant growth is not optimal during growth.
Table 3 shows that optimal bacterial growth did not occur at the start of the experiment (14 dap) because the bacteria had a lag phase that required adaptation for growth when moving to a new medium. The appearance of trifoliate leaves at 21 dap was caused by the collaboration of nitrogen-fixing and phosphate-solubilizing bacteria. The availability of nitrogen and phosphorus elements aids photosynthesis by producing more and more photosynthate, which is then transported to the plant's vegetative parts to form stems and leaves.
Table 4. Effect of indigenous bacterial on soybean on root length
|
Treatment |
Root length (cm) |
|
Without inoculation |
63.50 a |
|
NFB inoculation |
67.44 a |
|
PSB inoculation |
62.70 a |
|
ODB inoculation |
60.70 a |
|
NFB+PSB+ODB |
64.70 a |
Note: The Duncan's Multiple Range Test at a 5% error level indicates that there is no statistically significant difference between the mean number and the letter that comes after it.
Table 5. Effect of indigenous bacterial on soybean fresh weight
|
Treatment |
Fresh weight (g) |
|
Without inoculation |
49.68 a |
|
NFB inoculation |
57.43 a |
|
PSB inoculation |
55.23 a |
|
ODB inoculation |
50.59 a |
|
NFB+PSB+ODB |
56.16 a |
Note: The Duncan's Multiple Range Test at a 5% error level indicates that there is no statistically significant difference between the mean number and the letter that comes after it.
Table 6. Effect of indigenous bacterial on soybean root to shoot ratio
|
Treatment |
Root to shoot ratio |
|
Without inoculation |
3.09 a |
|
NFB inoculation |
4.18 a |
|
PSB inoculation |
4.67 a |
|
ODB inoculation |
5.03 a |
|
NFB+PSB+ODB |
8.23 b |
Note: The Duncan's Multiple Range Test at a 5% error level indicates that there is no statistically significant difference between the mean number and the letter that comes after it.
Table 7. Effect of indigenous bacterial on soybean root nodule
|
Treatment |
Number of Root nodule |
|
|
Efective |
Non efective |
|
|
Without inoculation |
106,40 a |
81.40 a |
|
NFB inoculation |
104,40 a |
10.00 a |
|
PSB inoculation |
170,20 b |
37.40 a |
|
ODB inoculation |
168,20 b |
35.80 a |
|
NFB+PSB+ODB |
184,00 b |
45.80 a |
Note: The Duncan's Multiple Range Test at a 5% error level indicates that there is no statistically significant difference between the mean number and the letter that comes after it.
Low nutrient content in the growing media, sub-optimal soil bacterial activity and genetic factors in soybean plants are assumed because the number of leaves was not significantly different. The result following Hindersah and Matheus [29] stated that the application of biofertilizers containing Azotobacter chroococcum, Azotobacter vinelandii, Azospirillum sp., Acinetobacter sp., Pseudomonas cepacia, and Penicillium sp. as much as 20 mL/plant did not affect the number of leaves of mustard plants. According to the current study, an inoculant of up to 10 mL/polybag has yet to produce good results at 28 and 35 dap. The result is closely related to the number of bacterial cells introduced. The non-maximal growth of leaf area is most likely due to the use of inoculants that are not optimal, as it does not affect the number of leaves at 28 and 35 dap.
According to the findings, administering 10ml/polybag of inoculant with a density of 106 CFU/ml did not produce satisfactory results on the parameter of soybean root length (Table 4). The bacterial population is closely related to the density of bacterial cells. The more bacteria there are in the root area, the better bacteria can colonize the rhizosphere. We assumed that the bacteria inoculated on soybean growing media are still relatively low. Higher densities are required to allow the bacteria to associate optimally with the root area, allowing the roots to expand their root zone and absorb nutrients and water properly.
The fresh weight of the crop is proportional to the accumulation of photosynthetic results and the plant's water content. The water absorption by the plants affects the wet weight of the crop produced, so the roots play a role in increasing the wet weight of the stover.
Plant growth is related to the availability of nutrients and water in the soil, which are absorbed by the roots and can affect the fresh weight of the crop produced. Potential bacteria that play a role in plant roots will increase the supply of nutrients in the soil, ensuring that the activity of supplying nutrients required for photosynthesis is adequately fulfilled [30, 31].
Based on the observations showed on Table 5, any bacterial inoculant has yet to produce good growth on the crop's fresh weight. It must be linked to soil conditions that are inadequate for bacterial growth. Bacterial growth is specific and is influenced by pH, C-organic content, temperature, and water availability. It is suspected that the non-maximal fresh weight of the crop indicates that water availability in plant tissues may be more optimal, even supported by the lack of bacteria colonizing the rhizosphere affects plant water and nutrient absorption.
Table 6 demonstrated that the inoculation of mixed bacteria NFB, PSB, and ODB significantly affected the root-to-shoot ratio. This finding is consistent with Sukmasari et al. [32], who found that applying a 30 kg/ha microbial consortium has the highest root loss ratio.
The presence of abundant microbes around plants causes chemical changes in the substrate's composition, such as reducing oxygen supply, changing pH, and others that affect the lives of other species. The root shoot ratio is the ratio of the dry weight of shoots to the dry weight of roots. According to Irwan et al. [33] a root-to-shoot ratio higher than one (high) indicates more plant growth toward shoots, whereas a root-to-shoot ratio less than one (low) indicates plant growth toward roots. The ideal root-shoot ratio for a food crop is three.
Because of adaptations to maintain root and crown balance, improved root growth will stimulate shoot growth. The inoculation of a bacterial mixture is believed to raise the microbial population. It can enhance the environment for plant growth, resulting in better outcomes for growth and biomass production.
Based on Table 7, it is clear that the bacterial inoculation treatment had a significant impact on the effective root nodules. It is suspected that a mixture of NFB, PSB, and ODB can symbiotically interact with legume plants, infecting plant roots and forming root nodules. This suggests that the functional root nodules formed can contribute to plant growth via N fixation.
According to the observation results, no treatment produced significantly different results on non-effective root nodules. It is assumed that by inoculating the bacteria, the bacteria will be able to optimize the growth of effective root nodules and thus influence the nitrogen fixation process. The lectin (phytohemagglutinin) produced by the legume root system influences the association of bacteria and plant roots that form root nodules. Legume plants produce lectins with different bacterial species specificities [34]. The effectiveness of nitrogen fixation will be determined by the suitability of the relationship between the bacterial strain and the soybean varieties. Bradyrhizobium was found in high abundance in nodules in the more efficient and specific symbiosis between soybean and Bradyrhizobium, which increased upon inoculation, according to Bender et al. [35]. The main Rhizobium symbiont, on the other hand, was less abundant in common bean nodules and did not increase with inoculation, which may explain the legume's frequently reported lack of response to elite strain inoculation. Co-inoculation with Azospirillum reduced the abundance of host-compatible rhizobia in nodules, most likely due to rhizosphere competition, but increased the abundance of host-compatible rhizobia in root microbiomes.
3.3 Yield result
Table 8 demonstrates that any particular type of bacteria did not significantly impact the number of soybean pods, indicating that when the plants reached the generative phase, the inoculant administration at planting, 14 days after planting, and 35 days after planting was unable to adjust and carry out activities to the best of its ability. This result will impact the supply of nutrients and the photosynthesis process necessary for developing pods. According to Gardner et al. [36], a plant's genetic potential is another element that influences the number of pods it produces. Genetic features in plants can influence the number of pods they produce. Plants' genetic characteristics and nutrient adequacy largely determine the formation and filling of pods. Therefore, the pods formed in plants will be filled with photosynthates, forming seeds.
Kalay and Hindersah [37] researched the inoculation of phosphate-dissolving bacteria and found that doing so increases yields because phosphate-solubilizing bacteria can increase soil phosphate availability through the mechanisms of phosphate dissolution by organic acids or phosphate degradation by secreted phosphatases. P is the second-most significant element after N, and it is crucial for photosynthesis, root growth, and the production of flowers, fruits, and seeds [38]. Similarly, previous study found that the administration of P fertilizer significantly impacted the number of filled pods per plant of mungbean [39] and cowpea [40].
The lack of available nutrients at the time of filling the seeds causes the translocation of assimilate to the seeds to decrease, and as a result, the filled pods need to be optimally filled. It is believed that the seed-filling process is being hindered since the contact between bacteria and plants have started to decline, the bacterial population has reduced, and the C-organic content has diminished. One factor affecting soybean productivity is the number of soybean seeds. The production of soybeans increases as more seeds are produced. Variations in the number of blooms present at the time of flower creation and the frequency of reproductive organ miscarriage affect the number of seeds produced by each plant. Therefore, the primary determinant of yield is the number of pods on the plant. The current study adopts Mimbar's [41] theory, which holds that fertilization-i.e., when pollen fertilizes the egg in the ovary-determines the number of seeds or pods. The weight of the photosynthate storage organs, particularly the yield organs (seeds), is low due to the low photosynthate received by plant organs. Seed weight per plant as a measure of yield in soybean farming is regarded by Salimi and Moradi [42] as a significant characteristic of soybean seed output.
During seed filling, both genetic and environmental variables affect each plant's individual seed size. During the seed-filling process, photosynthate produced by plants after flowering is translocated. The weight of the seed can be increased by photosynthate that is produced or stored during seed filling. Seed weight increase is the consequence of assimilation of remobilization from vegetative organs like leaves and stems to the seeds as well as photosynthesis outcomes that are directly directed to the seeds [43].
The weight of 100 dried seeds can determine the size of the seed endosperm. The main portion of the seed is the endosperm, which serves as a storage area for nourishment. In this experiment, the weight of 100 dried seeds was still less than the production potential, which ranged from 14 to 15 g, whereas the Devon-2 variety's potential maximum was 17.03 g (Table 9). As a result, the studied indigenous bacteria injection could have produced better outcomes. The bacterial inoculant utilized may also impact the soybean plants' weight limit of 100 seeds, which has not yet been reached. The efficiency of the interaction with the host plant cannot be explained because biochemical and physiological tests on the bacterial inoculant utilized have yet to be performed.
Table 8. The effect of indigenous bacterial inoculation on the number of pods per plant
|
Treatment |
Number of pods per plant |
|
without inoculation |
40.92 a |
|
NFB inoculation |
42.25 a |
|
PSB inoculation |
45.75 a |
|
ODB inoculation |
44.15 a |
|
NFB+PSB+ODB |
42.40 a |
Note: The Duncan's Multiple Range Test at a 5% error level indicates that there is no statistically significant difference between the mean number and the letter that comes after it.
Table 9. Effect of indigenous bacteria on the number of seeds, seed weight and weight of 100 soybean seeds
|
Jenis bakteri |
number of seeds / Plant |
Seed weight/ Plant (g) |
weight of 100 seeds (g) |
|
without inoculation |
90.22 a |
13.31 a |
15.10 a |
|
NFB inoculation |
94.40 a |
12.78 a |
15.11 a |
|
PSB inoculation |
101.26 a |
14.17 a |
14.47 a |
|
ODB inoculation |
97.28 a |
14.98 a |
15.37 a |
|
NFB+PSB+ODB |
94.15 a |
12.50 a |
14.85 a |
Note: The Duncan's Multiple Range Test at a 5% error level indicates that there is no statistically significant difference between the mean number and the letter that comes after it.
The total number of bacteria successfully isolated from the soil around the Mugarsari campus was 70, with 18 NFB isolates, 16 PSB isolates, and 36 ODB isolates. Seventeen bacteria isolate with potential that have passed qualitative and hypersensitive tests, including 4 NFB isolates, 8 PSB isolates, and 5 ODB isolates.
The application of indigenous bacterial isolates at a concentration of 10 mL/polybag (bacterial density 106 CFU/mL) affected the number of leaves, chlorophyll content, effective root nodules, and root to shoot ratio. However, it did not affect plant height, root length, number of non-effective root nodules, fresh weight of stover, number of pods per plant, number of seeds per plant, seed weight per plant, and weight of 100 dry soybean seeds Devon 2 variety. The number of bacteria administered to the plant media played a role in the outcome. The optimal number of colonies administered must be reviewed in order to determine the most optimal amount given. Inadequate organic sources for bacterial growth could have caused the non-maximum growth and yield of soybeans in this study resulting limited number of growing bacterial populations.
The most significant finding of this study was that adding local microbes can, on average, increase the growth and yield of soybeans. There are varying outcomes based on bacterial type for various growth and yield parameters. Different inoculants have positive effects on various parameters. Overall, however, combining three types of inoculants did not produce superior results than a single type. It suggests that the diversity of local microorganisms is not always associated with greater responses. Inoculants may be unable to acclimate well to the planting medium due to competition among them, thereby hindering the functional mechanism of bacteria in stimulating plant growth.
Overall, these findings suggest significant opportunities for future research on local strategies for sustainable integrated soil fertility management to increase soybean yield.
The authors would like to thank the LP2M-PMP of Universitas Siliwangi which has funded this research.
[1] FAO. (2018). Country factsheet small family farms: Indonesia. https://www.fao.org/3/i8881en/I8881EN.pdf, access on 21st May, 2023.
[2] Irawan, S., Antriyandarti, E. (2021). Fertilizer application, climate change and rice production in rural Java. In IOP Conference Series: Earth and Environmental Science, IOP Publishing, 755(1): 012086. https://doi.org/10.1088/1755-1315/755/1/012086
[3] Alta, A., Setiawan, I., Fauzi, A.N. (2021). Beyond fertilizer and seed subsidies: rethinking support to incentivize productivity and drive competition in agricultural input markets. CIPS Policy Papers, Center for Indonesian Policy Studies (CIPS), 43.
[4] García-Orenes, F., Morugán-Coronado, A., Zornoza, R., Cerdà, A., Scow, K. (2016). Correction: changes in soil microbial community structure influenced by agricultural management practices in a mediterranean agro-ecosystem. PLoS One, 11(3): e0152958. https://doi.org/10.1371/journal.pone.0152958
[5] Bertola, M., Ferrarini, A., Visioli, G. (2021). Improvement of soil microbial diversity through sustainable agricultural practices and its evaluation by-omics approaches: a perspective for the environment, food quality and human safety. Microorganisms, 9(7): 1400. https://doi.org/10.3390/microorganisms9071400
[6] Priyadi, R., Juhaeni, A.H., Meylani, V., Fudholi, A. (2022). The development of inorganic fertilizer and bio-fertilizer combination and the effectiveness of application on the growth and production of red chili. International Journal of Design & Nature and Ecodynamics, 17(1): 87-93. https://doi.org/10.18280/ijdne.170111
[7] Kumar, S., Sindhu, S.S., Kumar, R. (2022). Biofertilizers: an ecofriendly technology for nutrient recycling and environmental sustainability. Current Research in Microbial Sciences, 3: 100094. https://doi.org/10.1016/j.crmicr.2021.100094
[8] Batubara, U.M., Susilawati, I.O., Riany, H. (2015). Isolasi dan karakterisasi bakteri indigenous tanah di kawasan kampus Universitas Jambi. PROSIDING SEMIRATA 2015, 4(1).
[9] Ratrinia, P.W., Uju, P.S. (2017). Efektivitas penambahan bioaktivator laut dan limbah cair surimi pada karakteristik pupuk organik cair dari sargassum sp. Jurnal Pengolahan Hasil Perikanan Indonesia, 19(3): 309-320.
[10] Tang, A., Haruna, A.O., Majid, N.M.A., Jalloh, M.B. (2020). Potential PGPR properties of cellulolytic, nitrogen-fixing, phosphate-solubilizing bacteria in rehabilitated tropical forest soil. Microorganisms, 8(3): 442. https://doi.org/10.3390/microorganisms8030442
[11] Harsono, A., Harnowo, D., Ginting, E., Elisabeth, D.A.A. (2022). Soybean in Indonesia: current status, challenges and opportunities to achieve self-sufficiency. Legumes Research, 1. https://doi.org/10.5772/intechopen.101264
[12] Muslikah, S., Sudiarso, S., Setyobudi, L. (2016). Effect of inoculation and time of application of microbes on growth and yield of soybean (Glycine max (L.) Merrill). Journal of Degraded and Mining Lands Management, 4(1): 709-715. https://doi.org/10.15243/jdmlm.2016.041.709
[13] Pedrozo, A., Girelli de Oliveira, N.J., Alberton, O. (2018). Biological nitrogen fixation and agronomic features of soybean (Glycine max (L.) Merr.) crop under different doses of inoculant. Acta Agronómica, 67(2): 297-302. https://doi.org/10.15446/acag.v67n2.56375
[14] Cordeiro, C.F.D.S., Echer, F.R. (2019). Interactive effects of nitrogen-fixing bacteria inoculation and nitrogen fertilization on soybean yield in unfavorable edaphoclimatic environments. Scientific Reports, 9(1): 15606. https://doi.org/10.1038/s41598-019-52131-7
[15] Tikhonova, E.N., Menko, E.V., Kravchenko, I.K. (2019). Chapter 1-bacterial fertilizers for soybean plants: current status and future prospects. New and Future Developments in Microbial Biotechnology and Bioengineering, 1-20. https://doi.org/10.1016/B978-0-12-818258-1.00001-7
[16] Gitonga, N.M., Njeru, E.M., Cheruiyot, R., Maingi, J.M. (2021). Bradyrhizobium inoculation has a greater effect on soybean growth, production and yield quality in organic than conventional farming systems. Cogent Food & Agriculture, 7(1): 1935529. https://doi.org/10.1080/23311932.2021.1935529
[17] Jabborova, D., Kannepalli, A., Davranov, K., Narimanov, A., Enakiev, Y., Syed, A., Elgorban, A.M., Bahkali, A.H., Wirth, S., Sayyed, R.Z., Gafur, A. (2021). Co-inoculation of rhizobacteria promotes growth, yield, and nutrient contents in soybean and improves soil enzymes and nutrients under drought conditions. Scientific Reports, 11(1): 22081. https://doi.org/10.1038/s41598-021-01337-9
[18] Cappuccino, J.G., Sherman, N. (2014). Microbiology: A Laboratory Manual (10th edition). Pearson education inc.
[19] Zuraidah, Z. (2013). Pengujian beberapa bakteri penghambat pertumbuhan xanthomonas oryzae pv. oryzae Pada Tanaman Padi. Biologi Edukasi: Jurnal Ilmiah Pendidikan Biologi, 5(1): 18-24.
[20] Hindersah, R., Matheus, J. (2015). Respons pertumbuhan vegetatif jagung di tailing tambang timah terkontaminasi kadmium setelah inokulasi bakteri indogenus. Agrologia, 4(1). http://doi.org/10.30598/a.v4i1.211
[21] Dwidjoseputro, D. (2005). Dasar-dasar mikrobiologi. Djambatan: Jakarta.
[22] Nugroho, F.T., Setiawan, A.W. (2021). Isolasi dan karakterisasi bakteri pada tanah organik dan anorganik di kec. Kopeng dan kec. Magelang. Agriland: Jurnal Ilmu Pertanian, 9(1): 17-26. https://doi.org/10.30743/agr.v9i1.3931
[23] Leboffe, M.J., Pierce, B.E. (2012). Microbiology laboratory theory and application: second edition. Morton Publishing Companies.
[24] Schaad, N.W., Jones, J.B., Chun, W. (2001). Laboratory guide for identification of plant pathogenic bacteria, third edition. Plant Pathology, 50(6): 812-814. https://doi.org/10.1046/j.1365-3059.2001.00635.x
[25] Umesha, S., Richardson, P.A., Kong, P., Hong, C.X. (2008). A novel indicator plant to test the hypersensitivity of phytopathogenic bacteria. Journal of Microbiological Methods, 72(1): 95-97. https://doi.org/10.1016/j.mimet.2007.11.002
[26] García-Orenes, F., Morugán-Coronado, A., Zornoza, R., Scow, K. (2013). Changes in soil microbial community structure influenced by agricultural management practices in a Mediterranean agro-ecosystem. PloS one, 8(11): e80522. https://doi.org/10.1371/journal.pone.0080522
[27] Latupapua, H.J.D., Suliasih, S. (2018). Daya Pacu Mikroba Pelarut Fosfat dan Penambat Nitrogen pada Tanaman Jagung. Jurnal Biologi Indonesia, 3(2). https://doi.org/10.14203/jbi.v3i2.3477
[28] Glick, B.R. (2012). Plant growth-promoting bacteria: mechanisms and applications. Scientifica, 2012. https://doi.org/10.6064/2012/963401
[29] Hindersah, R., Matheus, J. (2015). Respons pertumbuhan vegetatif jagung di tailing tambang timah terkontaminasi kadmium setelah inokulasi bakteri indogenus. Agrologia, 4(1). http://doi.org/10.30598/a.v4i1.211
[30] Jacoby, R., Peukert, M., Succurro, A., Koprivova, A., Kopriva, S. (2017). The role of soil microorganisms in plant mineral nutrition-current knowledge and future directions. Frontiers in Plant Science, 8: 1617. https://doi.org/10.3389/fpls.2017.01617
[31] Khalil, H.P.S.A., Hossain, M.S., Rosamah, E., Azli, N.A., Saddon, N., Davoudpoura, Y., Islam, M.N., Dungani, R. (2015). The role of soil properties and it’s interaction towards quality plant fiber: a review. Renewable and Sustainable Energy Reviews, 43: 1006-1015. https://doi.org/10.1016/j.rser.2014.11.099
[32] Sukmasari, M.D., Atma, A., dan Indra Herdiana, W. (2018). Pertumbuhan tanaman kedelai (Glycine max L. Merrill) dengan pemberian pupuk hayati konsorsium dan fungi mikoriza arbuskular. Prosiding Konser Karya Ilmiah Tingkat Nasional Tahun 2018, 77-84.
[33] Irwan, A.W., Nurmala, T., Nira, T.D. (2017). Pengaruh jarak tanam berbeda dan berbagai dosis pupuk kandang ayam terhadap pertumbuhan dan hasil tanaman hanjeli pulut (Coix lacryma-jobi L.) di dataran tinggi Punclut. Kultivasi, 16(1): 233-245. https://doi.org/10.24198/kultivasi.v16i1.11719
[34] Lagarda-Diaz, I., Guzman-Partida, A.M., Vazquez-Moreno, L. (2017). Legume lectins: proteins with diverse applications. International Journal of Molecular Sciences, 18(6): 1242. https://doi.org/10.3390/ijms18061242
[35] Bender, F.R., Alves, L.C., da Silva, J.F.M., Ribeiro, R.A., Pauli, G., Nogueira, M.A., Hungria, M. (2022). Microbiome of nodules and roots of soybean and common bean: searching for differences associated with contrasting performances in symbiotic nitrogen fixation. International Journal of Molecular Sciences, 23(19): 12035. https://doi.org/10.3390/ijms231912035
[36] Gardner, F.P., Pearce, R.B., Mitchell, R.L. (1991). Physiology of crop plants (fisiologi tanaman budi daya, alih bahasa oleh susilo). Jakarta. University of Indonesia.
[37] Kalay, A.M., Hindersah, R. (2016). Efek pemberian pupuk hayati konsorsium terhadap pertumbuhan dan hasil tanaman sawi (Brassica juncea L.). Jurnal Agroekoteknologi, 8(2): 131-138. http://doi.org/10.33512/j.agrtek.v8i2.1486
[38] Simanungkalit, R.D.M., Suriadikarta, D.A., Saraswati, R., Setyorini, D., Hartatik, W. (2006). Pupuk organik dan pupuk hayati, balai besar penelitian dan pengembangan sumberdaya lahan pertanian. Balai Penelitian dan Pengembangan Pertanian.
[39] Nasution, F.M., Hasanah, Y., Mariati, M. (2020). Production response of mung bean (Vigna radiata L.) on the application of phosphorus fertilizer and oil palm bunch ash. Indonesian Journal of Agricultural Research, 3(1): 48-55. https://doi.org/10.32734/injar.v3i1.3839
[40] Singh, A., Baoule, A.L., Ahmed, H.G., Dikko, A.U., Aliyu, U., Sokoto, M.B., Alhassn, J., Musa, M., Haliru, B. (2011). Influence of phosphorus on the performance of cowpea (vigna unguiculata (l) walp.) Varieties in the sudan savanna of nigeria. Agricultural Sciences, 2(03): 313-317. https://doi.org/10.4236/as.2011.23042
[41] Muhuria, L. (2007). Mekanisme fisiologi dan pewarisan sifat toleransi tanaman kedelai (Glycine max (L) Merrill) terhadap intensitas cahaya rendah. Disertasi, Sekolah Pascasarjana, IPB, Bogor.
[42] Salimi, S., Moradi, S. (2012). Effect the correlation, regression and path analysis in soybean genotypes (Glycine max L.) under moisture and normal condition. International Journal of Agronomy and Plant Production, 3(10): 447-454.
[43] Schiltz, S., Munier-Jolain, N., Jeudy, C., Burstin, J., Salon, C. (2005). Dynamics of exogenous nitrogen partitioning and nitrogen remobilization from vegetative organs in pea revealed by 15N in vivo labeling throughout seed filling. Plant Physiology, 137(4): 1463-1473. https://doi.org/10.1104/pp.104.056713