Comparison of Bioethanol Production Using Saccharomyces cerevisiae and Zymomonas mobilis in Fermented Jackfruit Peel Treated with Blend Crude Cellulose Enzymes

Trianik Widyaningrum* | Listiatie Budi Utami | Indro Prastowo | Vita Meylani | Adi Permadi
© 2022 IIETA. This article is published by IIETA and is licensed under the CC BY 4.0 license (http://creativecommons.org/licenses/by/4.0/).
OPEN ACCESS
Bioethanol is an alternative energy source that is increasingly needed along with the depletion of petroleum stocks. Bioethanol can be produced by utilizing various wastes, including jackfruit peel and a mixture of crude cellulose enzymes from Trichoderma reesei and Aspergillus niger as well as fermentation using Saccharomyces cerevisiae and Zymomonas mobilis. This study aimed to compare the production of bioethanol using S. cerevisiae and Z. mobilis in fermented jackfruit peel with a mixture of crude cellulose enzymes from T. reesei and A. niger. The experimental design used a completely randomized design with the ratio of crude cellulase enzymes from T. reesei and A. niger as independent variables (0:0), (1:0), (0:1), (1:1), (1:2), (2:1), (1:3), and (3:1) as well as sugar and ethanol content as dependent variables. The sugar content was determined using the DNS method, while the ethanol content was determined using an alcoholmeter. Data analysis used one way ANOVA assisted by SPSS 16. The results showed that the highest sugar content (14.21 percent) was obtained in the ratio of crude cellulase enzymes T. reesei and A. niger (1:3), while the highest ethanol content (3, 16 percent) at a ratio of 1: 2 and fermented using Z. mobilis.
blend cellulase enzyme, jackfruit peel, Saccharomyces cerevisiae, Zymomonas mobilis
Fossil fuels, primarily oil, coal, and natural gas, are the primary energy source for most businesses and remain the world's primary raw materials for energy production. Currently, fossil fuels dominate the international energy industry, worth approximately 1.5 trillion dollars [1]. These resources, however, are no longer considered sustainable, and their availability has decreased significantly [2]. These fuels have negative environmental consequences, such as global warming due to greenhouse gas emissions [3]. As a result, alternative energy sources that are renewable, sustainable, and ecologically benign, such as bioethanol, are required [4].
Bioethanol can be produced from cellulose and glucose-based biomass, such as starch-containing biomass (corn, cassava, and sorghum), sugar-containing biomass (molasses and sap), and cellulose-containing biomass (cassava, rice straw, bagasse, and jackfruit peel) [5]. Jackfruit peel is still underutilized and ends up as waste. Jackfruit peel includes 15.87 percent carbs, 38.69 percent cellulose, and 1.30 percent protein [6]. Typically discarded, the jackfruit's peel has the potential to be used to produce bioethanol. Jackfruit peel contains approximately 14.74 percent sugar, making it suitable for use as bioethanol [7]. Cellulolytic microorganisms capable of producing cellulase enzymes, such as Trichoderma reesai and Aspergillus niger, are required to produce bioethanol from cellulose-containing materials. Trichoderma reesei can degrade cellulose and starch to glucose by the production of cellulolytic enzymes, specifically endoglucanase and exoglucanase, which hydrolyze cellulose [8]. Trichoderma reesei may also manufacture endo-1,4-glucanase and exo-1,4-glucanase enzymes in quantities up to 80%, albeit at a lower rate than Aspergillus niger [9]. Aspergillus niger is capable of producing large amounts of -glucosidase, as well as endo-1,4-glucanase and exo-1,4-glucanase. Additionally, exo-1,4-glucanase will cleave the cellulose chain's ends and generate cellobiose. The resulting cellobiose will be converted to glucose molecules by -glucosidase [9]. The conversion of cellulose by T. reesei and A. niger produces glucose, which can employ in the fermentation process to make bioethanol. Microorganisms such as Saccharomyces cerevisiae and Zymomonas mobilis carry out the sugar fermentation process. S. cerevisiae offers several advantages over other microbes used in the bioethanol production process, including being more ecologically friendly, versatile, easy to procure, and resistant to high alcohol content interactions [10]. Zymomonas mobilis ferments more specifically than yeast [11]. This study aimed to compare the amounts of bioethanol fermented jackfruit peel to crude enzymes T. reesei and A. niger after ratio treatment.
2.1 Preparation sample
On Jl. Ipda Tut Harsono, in the Umbulharjo sub-district of Yogyakarta, jackfruit peel is processed to be powered. The Food & Nutrition Culture Collection (FNCC) at the University of Georgia provided cultures of Trichoderma reesei, Aspergillus niger, Saccharomyces cerevisiae, and Zymomonas mobilis.
Trichoderma reesei and Aspergillus niger strains were cultivated on PDA (with ingredients 5 g of PDA powder, and 100 mL of distilled water) and cultured for seven days at room temperature [12]. Zymomonas mobilis cultures were cultivated on NA medium (0.42 g of NA media and 15 mL of distilled water) and incubated at room temperature for 24 hours [13].
2.2 Preparation of nutrient solutions
3 g urea powder per liter, 10 g (NH4)2SO4 powder per liter, 3 g KH2PO4 powder per liter, 0.5 g MgSO4 powder per liter 7H2O powder, 0.5 g/L calcium chloride H2O L employs an analytical balance. Combine all of the above components in a 200 mL beaker glass and top with 100 mL aquadest. All ingredients were thoroughly mixed using a sterilized glass stirrer [14].
2.3 Production of the cellulase enzyme
Each 250 mL Erlenmeyer was filled with 5 grams of jackfruit peel powder and sealed with 25 mL of nutrient solution. Sterilization of the mixed medium Suspended A. niger cultures were cultivated at room temperature for eight days. In contrast, another Erlenmeyer was cultured for T. reesei and incubated for six days at room temperature. Each result of the incubation was added to 100 mL of an 80 percent tween solution and agitated at 150 rpm for 120 minutes at room temperature. Centrifuge the solution for ten minutes at 3000 rpm. Employed the resulting supernatant as a crude enzyme extract [14].
2.4 Jackfruit peel fermentation using crude enzymes
200 grams of jackfruit peel powder was placed in an Erlenmeyer, followed by 1,600 mL of distilled water (three replicates), and then brought to a boil. Filtration of the decoction separates the substrate from the cooking water. Divided the substrate into 48 100 mL fermentation bottles. The addition of crude enzymes T.reesei and A.niger at a concentration of up to 10% according to the treatment with variations (0:0); (1:0); (0,1), (1:1), (1:2), (2:1), (1:3), (3:1) using a measuring pipette aseptically three times (48 bottles of fermentation), then stirred with a sterile glass stirrer. Covering the fermentation bottles with sterile cotton and lining them with aluminum foil, incubating them at 37oC for 24 hours. The sugar content of the hydrolysis products was determined using the DNS technique [15].
2.5 Saccharomyces cerevisiae with Zymomonas amobilis treatment
Hydrolysis findings using crude enzymes (24 bottles) combined with a 10% culture of S. cerevisiae cultured for 72 hours [17]. Additional 24 bottles of fermentation + 10% culture of Zymomonas immobilis fermented for 96 hours [16]. Distilled the fermentation products of Saccharomyces cerevisiae and Zymomonas amobilis, and the resulting bioethanol is quantified using an alcohol meter [17].
3.1 Sugar levels in the peel of jackfruit before and after pretreatment with NaOH
Based on the research, we found various of the sugar levels in the peel of jackfruit before and after pretreatment with NaOH (Table 1).
Table 1. Jackfruit peel substrate sugar content
|
Replication |
Reducing sugar content |
|
|
|
Before Pretreatment (%) |
After Pretreatment (%) |
|
1 |
8.33 |
9.18 |
|
2 |
9.35 |
10.29 |
|
3 |
11.06 |
9.72 |
|
Average |
9.58 |
9.73 |
After crude enzyme treatment, the sugar content of jackfruit peel substrate is as Figure 1.
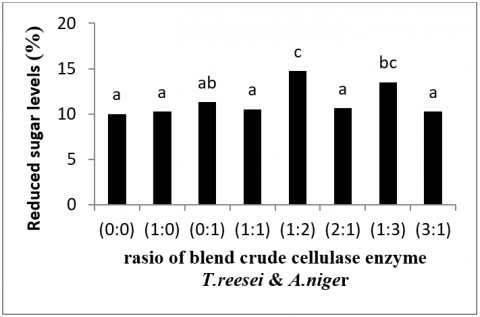
Figure 1. Reduced sugar levels after treatment with rasio of blend crude cellulase enzyme T. reesei & A. Niger
The same letter within each column do not differ significantly (p > 0.05) according to the Duncan test.
Sugar Content of Jackfruit Peel Following Fermentation with Saccharomyces cerevisiae in Figure 2.
Figure 2. Reduced sugar levels after treatment with ratio Saccharomyces cerevisiae
The same letter within each column do not differ significantly (p > 0.05) according to the Duncan test.
After Fermentation with S. cerevisiae, the ethanol content of jackfruit peel substrate was measured. Based on the results of the study, the measured ethanol content was as follows (Figure 3).
Figure 3. Bioethanol levels after treatment with Saccharomyces cerevisiae
The sugar content of jackfruit peel following treatment with Zymomonas mobilis is as follows (Figure 4).
Figure 4. Reduced sugar levels after treatment with Zymomonas mobilis
Bioethanol levels in jackfruit peel following treatment with Zymomonas mobilis is as follows (Figure 5).
The same letter within each column do not differ significantly (p > 0.05) according to the Duncan test.
Figure 5. Bioethanol levels after treatment with Zymomonas mobilis
4.1 Sugar levels in the peel of jackfruit before and after pretreatment with NaOH
Sugar content was determined before and after pretreatment to assess the ability of cellulose to be delignified/broken down, resulting in higher sugar content after pretreatment. According to Table 1, the outcomes of lowering sugar content before and after pretreatment were 9.58 percent and 9.73 percent, respectively. These findings indicate that the sugar concentration increased. Pretreatment removes lignin from lignocellulose and hydrolyses cellulose and hemicellulose into simple sugars to enhance the amount of sugar that will turn into bioethanol later [18].
4.2 Sugar levels after treatment with ratio of blend crude cellulase enzyme T. reesei & A. niger
According to the research findings, the sugar content of the jackfruit peelsubstrate after crude enzyme treatment ranges between 10.03 percent and 14.21 percent (Figure 1). The control sugar content was lower than the sugar content of all treatments. All crude enzyme treatments affected the sugar content since all calculation values were greater than the control sugar level. The treatment with the greatest sugar content, treatment 6, had a crude enzyme ratio of T. reesei: A. niger 1:3 and a sugar range of 14.21 percent. While treatment 1 had the lowest sugar content of all treatments, with a crude enzyme ratio of T. reesei: A. niger 1:0 of 10.40 percent. The crude T. reesei: A. niger 1:3 ratio produced the largest sugar content due to an increase in the activity of the crude enzyme Aspergillus niger, which is capable of producing high glucosidase [19]. The causal factor is decreased enzyme activity due to the lowest sugar content using the crude enzyme T. reesei: A. niger 1:0; The causative factor is the absence of crude enzymes from A. niger (0), so that only crude enzymes from T. reesei work so that the ability to convert cellulose into glucose is not optimal, besides that sugar levels also decrease due to glucose that has been hydrolyzed a lot so that sugar levels tend to be low. down or constant [19].
4.3 The Sugar Content of Jackfruit Peel Following Treatment with Saccharomyces Cerevisiae
According to the research results on the sugar content of fermented jackfruit peel substrate by Saccharomyces cerevisiae, shown in Figure 3, the average sugar content of fermented jackfruit peel substrate ranged between 10.02 percent and 14.73 percent. Treatment 4 had the greatest fermented sugar content, with a crude Trichoderma reesei: Aspergillus niger 1:2 ratio of 14.73 percent. Meanwhile, the lowest fermented sugar content was 10.27 percent when treated with a 1:0 ratio of crude enzyme T. reesei: A. niger. This is due to treatment 1 ratio of crude enzyme T. reesei: A. niger 1:0, crude enzyme from A. niger is not available (0), so that only crude enzyme from T. reesei works, conversion of cellulose to glucose is not optimal, resulting in low reducing sugar content (10.27%).
4.4 The sugar content of jackfruit peel following treatment with Zymomonas mobilis
According to the research results on the sugar content of fermented jackfruit peel substrate by Zymomonas mobilis, shown in Figure 4. The inclusion of 10 mL of Zymomonas mobilis altered the result of each treatment in terms of sugar reduction. The reducing sugar content produced after Z. mobilis treatment indicated that the highest reducing sugar content was 10.46 percent in the P2 treatment with a 1:1 ratio of A. niger to T. reesei and the lowest average sugar content was 8.44 percent in the P6 treatment with ratio of A. niger to T. reesei (1:3). Between pre- and post-fermentation with Z. mobilis, the sugar level reduced from 11.99 to 10.46 percent. The decreasing sugar levels the following fermentation was caused by Z. mobilis utilizing the sugar content of the fermentation medium to produce new cells and form ethanol [20]. According to Rahmadani et al. [21], Zymomonas mobilis eats sugar to generate bioethanol as the main metabolite. Apart from being transformed into bioethanol, sugar serves as a food source for bacteria to survive and reproduce.
4.5 Bioethanol levels in jackfruit peel following treatment with Zymomonas mobilis
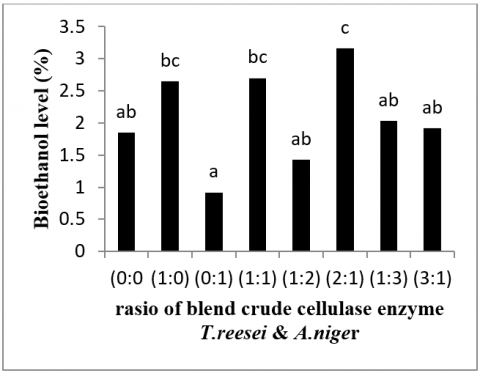
According to Figure 5, the ethanol content is between 1.04 percent and 1.32 percent. The greatest ethanol level was 1.32 percent in T. reesei and A. niger (1:1). Meanwhile, the control (T. reesei and A. niger 0:0) had the lowest ethanol concentration, at 1.04 percent. This finding is lower than that of a previous study [22, 23], which determined the ethanol content of jackfruit peel after treatment with water (2 percent). This yield was similarly lower than that of 25 grams of manganese seeds (4.78 percent) and 45 grams of manga seeds (1.48 percent) fermented with S. cerevisiae [24]. It could explain thelow ethanol content in this study because the longer the fermentation duration, the fewer microorganisms are created. As more bioethanol is formed, but fewer nutrients are available, the bioethanol produced is oxidized to carboxylic acid [20].
The maximum concentration of bioethanol was found in the crude enzyme A. niger: T. reesei (2: 1) ratio of 3.16 percent. In the ratio of crude enzyme A. niger: T. reesei, the lowest bioethanol content was 0.92 percent (1: 0). The average ethanol produced in this investigation was between 2-3%. The amount of bioethanol acquired in this study is greater than that achieved in previous research [23]; the ethanol content created from jackfruit peel is approximately 2% when yeast Saccharomyces cerevisiae is used. According to the study [25], the maximum amount of ethanol was produced using Saccharomyces cerevisiae, and the addition of 20% salt was 2%. It is consistent with the remark made by Fatimah et al. [26] that employing Z. mobilis bacterium is more successful than using Saccharomyces cerevisiae for creating higher ethanol levels.
The results of the measurement of the average pH after fermentation showed an increase from pH 7 to pH 8 (alkaline). The increase or decrease in the pH value was caused by A. niger and T. reesei cells not only consuming reducing sugars as nutrients for growth but also as producers of other products. Pangaribuan et al. [27] state that the increase in pH in a treatment can be caused by microorganisms, because microorganisms not only convert reducing sugars into bioethanol but also consume reducing sugars as nutrients to help metabolism. In addition, it can also produce organic acids so that it can increase or decrease pH. Meanwhile, according to Sadzvirani et al. [28, 29], this is because the enzyme has ampholytic properties, which means that the enzyme has a dissociation constant in the acid group and the base group.
This study indicates that the ratio of crude enzymes Aspergillus niger and Trichoderma reesei affects the sugar and bioethanol content of fermented jackfruit peel (Arthocharpus heterophyllus Lamk.) when Saccharomyces cerevisiae and Zymomonas mobilis are used. The maximum sugar concentration was 14.73 percent in the treatment of crude Trichoderma reesei: Aspergillus niger 1:2 ratio. The ratio of Aspergillus niger to Trichoderma reesei crude enzymes affects the bioethanol concentration of jackfruit peel. The greatest bioethanol concentration was 3.16 percent when crude enzyme (Aspergillus niger: Trichoderma reesei 2: 1) was fermented with Zymomonas mobilis. The bioethanol concentration of fermented jackfruit peel produced by Zymomonas mobilis was significantly higher than that produced by Saccharomyces cerevisiae.
The writers wish to express their gratitude to LPPM Ahmad Dahlan University. Contract Number: PD-174/SP3/LPPM-UAD/VI/2021.
[1] Gielen, D., Boshell, F., Saygin, D., Wagner, N., Gorini, R. (2019). The role of renewable energy in the global energy transformation. Energy Strategy Reviews, 24: 38-50. https://doi.org/10.1016/j.esr.2019.01.006
[2] Johnsson, F., Kjärstad, J., Rootzén, J. (2019). The threat to climate change mitigation posed by the abundance of fossil fuels. Climate Policy, 19(2): 258-274. https://doi.org/10.1080/14693062.2018.1483885
[3] Ahmed, W., Sarkar, B. (2018). Impact of carbon emissions in a sustainable supply chain management for a second generation biofuel. Journal of Cleaner Production, 186: 807-820. https://doi.org/10.1016/j.jclepro.2018.02.289
[4] Pelkmans, L., Van Dael, M., Junginger, M., et al. (2019). Long-term strategies for sustainable biomass imports in European bioenergy markets. Biofuels, Bioproducts and Biorefining, 13(2): 388-404. https://doi.org/10.1002/bbb.1857
[5] Said, M., dan Sri Mulyani Sabang, A.W.M. (2011). Synthesis of Bioetanol from Rice Straw (Oryza sativa L) By Fermentation. J. Akad. Kim., 3(4): 178-182.
[6] Hermawani, R.R., Ramadhani, D., Daya, A.M., Wahyudi, F., Sukemi, S. (2018). Nutrisi tepung kulit dan jerami nangka nutrition of jackfruit peel and straw flour. In Pendidikan Kimia FKIP UNMUL, pp. 76-79.
[7] Utami, R.K., Ganefati, S.P., Windarso, S.E. (2015). Pengaruh berat saccharomyces cereviceae dan waktu fermentasi kulit nangka terhadap kadar bioetanol yang dihasilkan. Jurnal Kesehatan Lingkungan, 6(4): 176-180.
[8] Ratanaphadit, K., Kaewjan, K., Palakas, S. (2010). Potential of glucoamylase and cellulase production using mixed culture of Aspergillus niger TISTR 3254 and Trichoderma reesei TISTR 3081. Culture, 15: 833-842.
[9] Kodri, Argo Dwi, B., Yulianingsih, R. (2013). Pemanfaatan enzim selulase dari Trichoderma reseei dan Aspergillus niger sebagai Katalisator Hidrolisis Enzimatik Jerami Padi dengan Pretreatment microwave. Jurnal Bioproses Komoditas Tropis, 1(1): 36-43.
[10] Azizah, N., Al-Baari, A., Mulyani, S. (2012). Pengaruh lama fermentasi terhadap kadar alkohol, pH, dan produksi gas pada proses fermentasi bioetanol dari whey dengan substitusi kulit nanas. Jurnal Aplikasi Teknologi Pangan, 1(2): 72-77. https://dx.doi.org/10.17728/JATP.V1I3.73
[11] Albert, Idiawati, N., Rudiansyah. (2015). Pembuatan bioetanol mengunakan Zymomonas mobilis dari limbah tongkol jagung. JKK: Jurnal Kimia Khatulistiwa, 4(2): 72-75.
[12] Fang, H., Zhao, C., Song, X.Y. (2010). Optimization of enzymatic hydrolysis of steam-exploded corn stover by two approaches: Response surface methodology or using cellulase from mixed cultures of Trichoderma reesei RUT-C30 and Aspergillus niger NL02. Bioresource technology, 101(11): 4111-4119. https://dx.doi.org/10.1016/j.biortech.2010.01.078
[13] Zhang, K., Feng, H. (2010). Fermentation potentials of Zymomonas mobilis and its application in ethanol production from low-cost raw sweet potato. African Journal of Biotechnology, 9(37): 6122-6128.
[14] Argo, B.D., Wardani, A.K., Zubaidah, E., Winarsih, S. (2014). The increasement of rice straw hydrolysis using Aspergillus niger. Research and Review in BioScience, 9(2): 41-44.
[15] Jackson, B., Jayanthy, T. (2014). Determination of sucrose in raw sugarcane juice by microwave method. Indian Journal of Science and Technology, 7(5): 566-570. https://dx.doi.org/10.17485/ijst/2014/v7i5.10
[16] He, M.X., Wu, B., Qin, H., et al. (2014). Zymomonas mobilis: A novel platform for future biorefineries. Biotechnology for Biofuels, 7: 101. https://doi.org/10.1186/1754-6834-7-101
[17] Widyaningrum, T., Parahadi, M. (2020). Bioethanol levels of dragon fruit (Hylocereus polyrhizus) peel with the addition of blend crude cellulase enzyme from Trichoderma reesei and Aspergillus niger. Journal of Tropical Biodiversity and Biotechnology, 5(1): 1. http://dx.doi.org/10.22146/jtbb.52189
[18] Hendrasarie, N., Mahendra, D.E. (2020). Pemanfaatan sampah sayur dari pasar tradisional untuk produksi bioetanol. Jurnal Serambi Engineering, 5(3): 1115-1122. http://dx.doi.org/10.32672/jse.v5i3.2075
[19] Oktavia, F.I., Argo, B.D., Lutfi, M. (2014). Enzymatic hydrolysis of bagasse utilizing enzymes cellulase from micro Trichoderma ressei and Aspergillus niger as catalyst in microwave pretreatment. Jurnal Keteknikan Pertanian Tropis dan Biosistem, 2(3): 256-262. http://dx.doi.org/10.1016/j.procbio.2011.01.022
[20] Irvan, Putri, A.W., Surbakti, S.U., Trisakti, B. (2016). Pengaruh konsentrasi ragi dan waktu fermentasi pada pembuatan bioetanol dari biji cempedak (Artocarpus champeden spreng). Jurnal Teknik Kimia USU, 5(2): 21-26. https://doi.org/10.32734/jtk.v5i2.1530
[21] Rahmadani, S., Muria, S.R., Utami, S.P. (2017). Produksi bioetanol dari mahkota nanas menggunakan bakteri Zymomonas mobilis dengan variasi konsentrasi inokulum dan penambahan nutrisi. Fteknik, 4(2): 1639-1642.
[22] Masturi, M., Cristina, A., Istiana, N., Sunarno, S., Dwijananti, P. (2017). Ethanol production from fermentation of arum Manis mango seeds (Mangifera Indica L.) using Saccharomyces crevisiaea. Jurnal Bahan Alam Terbarukan, 6(1): 56-60. https://doi.org/10.15294/jbat.v6i1.8139
[23] Rifa’i, M., Mukti, B.H., Lagiono (2018). Pengaruh perbedaan media air terhadap karakteristik hasil fermentasi kulit nangka. Jurnal Pendidikan Hayati, 4(2): 77-84.
[24] Meyrinta, K.A., Putri, R.D., Fatoni, R. (2018). Pembuatan bioetanol dari Jerami Nangka dengan Metode Fermentasi Menggunakan Saccharomyces cerevisiae. Jurnal Integrasi Proses, 7(1): 32-38. http://dx.doi.org/10.36055/jip.v7i1.2893
[25] Ulimaz, A. (2021). Kandungan etanol kulit nangka (Arthocharpus Heterophyllus Lamk). Nusantara Hasana Journal, 1(4): 1-6.
[26] Fatimah, Ginting, D., Sirait, V. (2017). Kinerja Mikroba Zymomonas mobilis dan Saccharomyces cerevisiae untuk menguraikan hidrolisat tongkol jagung menjadi bioetanol dengan pengaruh waktu fermentasi dan rasio penambahan mikroba. Jurnal Teknik Kimia USU, 6(2): 1-6. https://doi.org/10.32734/jtk.v6i2.1575
[27] Pangaribuan, R.N., Tambunan, G.A., Martgrita, M.M., Manurung, A. (2021). Potensi Kulit Buah untuk menghasilkan bioetanol dengan mengkaji kondisi, substrat, dan metode fermentasi. Journal of Applied Technology and Informatics Indonesia, 1(1): 433-470. https://doi.org/10.54074/jati.v1i1.7
[28] Sadzvirani, S., Restuhadi, F., Rossi, E. (2017) Bioethanol fermentation from very high gravity NYPA sap in batch fermentation with the addition of cordyceps mycelium powder and urea. Applied Science and Technology, 4(1): 1-15.
[29] Effendy, Antara, M., Muhardi, Pellokila, M.R., Mulyo, J.H. (2021). Identification of factors affecting decisions to adopt pesticides at lowland rice farms in Indonesia. International Journal of Design & Nature and Ecodynamics, 16(6): 717-723. https://doi.org/10.18280/ijdne.160614