Yusran Yusran* | Erniwati Erniwati | Akhmad Khumaidi | Ramadanil Pitopang | Rahmawati Rahmawati | I Nengah Korja | Elhayat Labiro | Rukmi Rukmi | Muthmainnah Muthmainnah | Haerani Maksum | Ratna Setyawati
© 2022 IIETA. This article is published by IIETA and is licensed under the CC BY 4.0 license (http://creativecommons.org/licenses/by/4.0/).
OPEN ACCESS
Lore Lindu National Park (LLNP) is one of the most important ecosystem protection areas in Indonesia, where there is a wide variety of ecosystems. Currently, mushroom diversity in LLNP is rarely reported. This study aims to determine the diversity of macro fungi in several types of land use in Toro village around Lore Lindu National Park, Central Sulawesi and its potential as a raw material for food and medicine. The research was conducted in primary forest (Wana Ngkiki and Wana), secondary forest (Pangale), shrubs (Oma) and agroforestry land (Pampa). A total of 148 species of macro mushrooms were found, which were distributed as many as 52 species in primary forest (Wana Ngkiki), 48 species in primary forest (Wana), 40 species in secondary forest (Pangale), 34 species in shrubs (Oma) and 50 species. in agroforestry land (Pampa). Furthermore, as many as 12 types of macro mushrooms are used as food and medicine by local indigenous people, namely: Pleurotus sp, Auricularia sp, Auricularia auricula-judae, Termytomyces sp, Lentinus sp, Schizophyllum commune, Agaricus subrutilescens, Boletus sp, Daldinia sp and Ganoderma lucidum. The number of macro-fungus species found in this area is certainly not a final investigation. Therefore, further expeditions to explore the diversity of macro fungi should be carried out in all parts of the park and in different seasons every year in order to obtain more complete data of macro fungi in the park.
diversity, indigenous peoples, land use types, Lore Lindu National Park, macro fungi
Lore Lindu National Park is one of the most important protected areas in Indonesia, and in 1977 it was declared by UNESCO as a biosphere reserve [1]. This area has conservation value and watershed protection for several major rivers in Central Sulawesi, Indonesia [2]. Ramadanil [3] reported that the main tree species in the park are Palaquium quercifollium, Palaquium obovatum, Castanopsis accuminatisima, Lithocarpus celebecus, Bischoffia javanica, Octomeles sumatrana, Pangium edule, Pterospermum celebicum, Aglaia argentea, Chionanthus ramiflorus and Polyscias nodosa. The condition of the forest in this area is still less disturbed by human activities, although in recent years, in the area there have been changes in land cover and land use, where some parts of the park have been converted to agricultural land. The long-term impact of these land use changes has resulted in a decline in the biodiversity population in this area. Furthermore, the selection of land use practices that are suitable for biodiversity conservation and the environment is very much needed, and of course research is needed on different aspects of biodiversity including microbial populations [4]. Crops provide a habitat for a wide and diverse community of macro fungi.
Macro mushrooms are a group of fungi that have macroscopic fruiting bodies, especially from Phyllum Ascomycota and Basidiomycota, so they can be easily seen with the naked eye. Macro fungi have a short reproductive stage in their life cycle [5]. They grow in areas with high humidity and water availability, where the rainy season is the most appropriate time for macro mushroom collection [6]. Many types of mushrooms in the world that are yet to be described. Estimates by Hawksworth & Lűcking [7] that there are about 2.2 – 3.8 million types of fungi in the world, and only about 6.7% have been described [8]. This number differs from the estimate by Mueller et al. [9], who reported that the total number of fungal species was only about 712,000 types. Of this total, Chang and Miles [10] reported that about 140,000 species were classified as macro fungi.
In tropical forest ecosystems, the total number of described fungal species is still very poor and unclear [8, 11, 12]. According to Canon et al. [13] that currently more than 93% of mushroom species in the world are still unknown. In Indonesia, it is estimated that around 200,000 species of fungi have been found [14], while in Malaysia, 70-80% of fungi have not been found [15, 16]. Specifically for macro mushrooms, Mueller et al. [9] estimated that the number of macro-fungal species in Tropical Asia ranged from 10,000 to 25,000. Furthermore, Desjardin et al. [17] estimated that some 300 species of the class basidiomycetes in Thailand represent less than 20% of the actual diversity of these macro fungi. While in Malaysia, Mueller et al. [9] reported that there are about 400 species members of Agaricus and Boletus groups were founded.
Basidiomycetes are a class of fungi represented by more than 20,000 species worldwide, including agarics, polypores, club and coral fungi, chantereles, jelly fungi, puffballs and tooth fungi, and yeasts. They play an important role in ecosystem functions and have various roles in terms of providing nutrients to plants, as pathogens, saprophytes and mutualism [17]. In addition to these functions, many types of macro fungi are a source of food and medicine [18-21] and also have potential as bioherbicides [22, 23]. The diversity of basidiomycetes in tropical countries has received less attention, including in Southeast Asia.
The indigenous people of Toro village are part of the Kaili tribe, an indigenous people living in the western part of the enclave of Lore Lindu National Park, a UNESCO biosphere reserve located on the island of Sulawesi. They have local wisdom in managing the environment by dividing the land management system based on the intensity of its use. Of the 22,300 hectares of traditional land belonging to the indigenous Toro community, 18,000 hectares are within the National Park and have been legally recognized by park authorities [24]. Ramadanil [3] indicates that the community of Toro divides its region in some zones, according to its function; is zone of primary forest, secondary forest, shrubs and agroforestry.
Data on the diversity of macro fungi in different land use types are important for planning and managing the biodiversity of an ecosystem. In Indonesia, research on biodiversity has been widely carried out, including on the island of Sulawesi. In the province of Central Sulawesi, research on biodiversity has been carried out especially on the diversity of plant species, including bryophytes and lichens, amphibians, reptiles, insects, bats, ants, birds and butterflies [3, 25-33]. However, research on the diversity of macro fungi in the area is still lacking. And specifically research on the diversity of macro fungi, has been carried out by Yusran et al., [34], where they found that the diversity of macro mushrooms was strongly influenced by different altitude levels from sea level in Lore Lindu National Park, Central Sulawesi, and several types of macro mushrooms were used by local communities as food and drug. Furthermore, research on the diversity of macro-mushroom species and their potential use in this area still needs to be done, considering the potential for macro-mushroom species, their role and use by the community as food or medicine in the future. This study aims to determine the diversity of macro mushrooms in several types of land use around Lore Lindu National Park, Central Sulawesi, Indonesia and their potential as food and medicine.
2.1 Research locations
The research was carried out on several types of land use around Toro village, South Kulawi sub-district, Sigi district. (longitude 01º22'52''-01º31'4'' S; latitude 120º1'37''-120º3'5'' E [35]. This village is located on the western border of Lore Lindu National Park, and administratively belongs to two districts, namely Sigi Regency and Poso Regency (Figure 1). The climate at the study site is a humid tropical climate characterized by high rainfall and high humidity.The average annual rainfall at the study site is between 1500 mm to 3000 mm, the average relative humidity is 85.17% and the average monthly temperature is 23.4℃ [36]. The research location is ± 100 km south of the city of Palu, the capital city of Central Sulawesi Province, Indonesia. In the study area, on the outskirts of the national park, it is characterized by primary forest, lightly disturbed primary forest to heavily disturbed, secondary forest and several land use systems with the main crops being cocoa, coffee, corn and rice [37]. The elevation of the selected site is between 800 to 1100 m above sea level, therefore it covers the range of elevations included in the sub-montane forest zone [38].
The collection of macro mushrooms was carried out on five types of land use with different intensity of use based on the local wisdom of indigenous peoples in Toro village, namely primary forest (Wana Ngkiki and Wana), secondary forest (Pangale), shrubs (Oma) and agroforestry (Pampa). The classification of land use types is presented in Table 1 [39].
Table 1. Overview of the characteristics of each type of land use as the research location in around Lore Lindu National Park
|
No |
Land Use Type/ Coordinates |
Characteristics |
|
1.
|
Primary Forest (Wana Ngkiki) S 01º 27'28.0” and E 120º 03'15.3” / |
Primary Forest, in the form of a primary forest area located on the top of the highest mountain, which is not allowed to have human activities and is located far from the village. This area is dominated by thick moss on the ground, there are also large trees with moss, and a little grass, several types of birds, as well as sources of clean air and storage of clean water reserves. (water reservoir). |
|
2. |
Primary Forest (Wana) S 01º 27'49.4” and E 120º 02'40.6”
|
Primary forest whose area is below Wana Ngkiki, is a large habitat for protected animals such as anoa, deer pigs, various types of animals. This area is a water catchment area when it rains, with a low intensity of use and very strict, it is forbidden to clear land or cut down trees. However, it is allowed to collect forest products such as forest, such as rattan, resins, medicines and fragrances. |
|
3. |
Secondary forest (Pangale) S 01º 29'24.2” and E 120º 02'08.2” |
Forest Secondary, is a transition between primary and secondary forest. Most of the forest area was cleared by residents and then abandoned decades ago, then experienced a succession similar to primary forest. In this zone, it is allowed to cut wood to build houses through rules or regulations that have been agreed upon by the community and customary institutions. Pangale is also used to collect rattan and wood for household and household purposes, Pandanus sp. for making mats and baskets, medicinal ingredients, resinous resin and fragrances. |
|
4. |
Shrubs (Oma) S 01º 29'22.4” and E 120º2'00.0” |
Ex-garden land that is intentionally left to be reprocessed for a certain period of time. Oma is also an area overgrown with medicinal plant species as well as a habitat for small birds and insects |
|
5. |
Agroforestry (Pampa) S 01º 29'30.3” and E 120º01'58.2” |
Agroforestry land where cacao and coffee are cultivated under the natural shade of large, unlogged trees on the outskirts of the forest. |
Figure 1. Map of research locations in Toro Village, around Lore Lindu National Park, Central Sulawesi, Indonesia (red box)
2.2 Survey, inventory and identification of macro fungus species
Inventory of mushrooms was done by sampling method intentionally (purposive sampling). Plot size and fruiting body sampling of macro fungal species were designed as suggested by Engola et al. [40]. Each type of land use was sampled in plots of different sizes. Two transects were made on the agroforestry (Pampa) land. Each transect consists of ten plots measuring 50x20 m (0.1 ha), located to the left and right of the 500m long transect. The distance between plots along the transect was 100m, and the distance between plots on the left and right transects was 10m. In forest areas (Wana Ngkiki, Wana, Pangale and Oma), ten strip plots measuring 5x200 m (0.1 ha) were used to facilitate the collection of macro mushroom fruiting bodies. Plots were selected as much as possible at the same height for each land use type to avoid differences in fungal composition caused by macroclimate. A total of 40 plots were made at the research location above.
All macro fungal sporocarps in the observation plot were collected both those that grew as saprophytes on litter, rotting wood and soil as well as those that were parasites on trees, as well as ectomycorrhizal fungi. The soil attached to the sporocarp was cleaned carefully, then the sporocarp was stored in a paper bag. The morphology of the sporocarps and their habitat conditions were recorded. Furthermore, the microscopic characteristics of macro fungi such as the shape and color of the spores were observed using a microscope. Mushroom abundance (a measure of the total number of fungi) was determined by manual counting of macrofungi and/or their fruiting bodies in the mapped area. The fruiting bodies of macro mushrooms that grew solitary, scattered, or in groups were all recorded as single observations without counting the number of sporocarps [41, 42]. Fresh specimens were photographed in the field.
The macro mushroom sporocarps were then dried using an electric fruit dryer at 40℃ for 15-20 hours and stored in a sealed plastic bag with silica gel at room temperature. The mushroom samples were stored in the Laboratory of Forestry Sciences, Faculty of Forestry, Tadulako University, Palu, Central Sulawesi. Macro fungi were identified as morphospecies based on macro and micro morphological characteristics, using the manual books and monograph by the references [17, 43-52]. The nomenclature of macro mushrooms follows Index Fungorum [53].
The data on the Ethnomycology of macro mushrooms found was carried out by interviewing several members of the local community, especially those who work as traditional healers, village heads, community leaders and parents (elders) who have experience in using macro mushrooms as food or medicine. The information asked related to customary knowledge, beliefs and practices of the use of these macro mushrooms.
2.3 Data analysis
The Shannon-Wiener diversity index of macro fungi was calculated as proposed by Margalef [54].
$H=\sum(n / N) \log _{c}(n / N)$ (1)
where, H is the diversity index, N is the total number of individuals of all species and n is the total number of individuals of the individual species.
The Simpson diversity index was calculated as proposed by Simpson [55].
Simpson Index of Diversity = 1-D,
$\mathrm{D}=\frac{\sum n(n-1)}{N(N-1)}$ (2)
where, n is the total number of organisms of a given species, N is the total number of organisms of all species and D is the Simpson's Index.
Based on the diversity index value, evenness of macro fungi was also calculated as proposed by Pielou [56].
$e=H / \log \mathrm{S}$ (3)
where, e is evenness, H is the Shannon diversity index and S is the number of species.
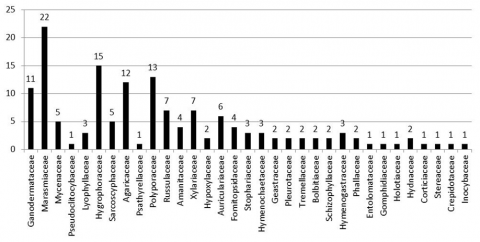
The results showed that there were 148 species of macro mushrooms divided into 33 families which were collected from several types of land use around Lore Lindu National Park, Central Sulawesi. The eleven types of macro mushrooms belong to the Ganodermataceae family, twenty-two belong to the Marasmiaceae family, fifteen to the Hygrophoraceae family, the Mycenaceae and Sarcoscyphaceae families each in the lime genus, twelve to the Agaricaceae family, thirteen to the Polyporaceae family, seven to the Russulacea family, seven to the family Xylariaceae, six to the family Auriculariaceae, the family Fomitopsidaceae and Amanitaceae each consisting of four genera, family Stophariaceae, Lyophyllaceae, Hymenochaetaceae and Hymenogastraceae each three genera, family Hypoxylaceae, Geastraceae, Pleurotaceae, Tremellaceae, Bolbitiaceae, Schizophyllaceae, Phallaceae and Hydnaceae each consist of 2 species. And the rest of the families Pseudoclitocybaceae, Psathyrellaceae, Entolomataceae, Gomphidiaceae, Holotiaceae, Corticiaceae, Stereaceae, Crepidotaceae and Inocybaceae, each represented by only one species (Figure 2). All of the macro-fungal families above belong to the phylum Basidiomycetes, while the other three macro-fungal families namely Sarcoscyphaceae, Xylariaceae and Holotiaceae are included in the phyllum Ascomycetes (Table 2 and Figure 2).
Table 2. Distribution of types and families of macro fungi in several types of land use in the vicinity Lore Lindu National Park
|
No |
Family |
Type |
Total |
|
1 |
Ganodermataceae |
Ganoderma applanatum (Press.) Pat, Ganoderma lucidum (Curtis) P. Karst., Ganoderma chiungchungense XLWu., Ganoderma sp.1, Ganoderma sp.2, Ganoderma sp.3, Ganoderma sp.4, Ganoderma sp.5, Ganoderma sp.6, Amauroderma rugonsum (Blume & T. Nees) Torrend, Amauroderma sp. |
11 |
|
2 |
Marasmiaceae |
Crinipellis stipitaria (Fr.) Pat., Crinipellis zonata (Peck) Sacc., Campanella junghuhnii (Mont.) Singer, Marasmius maximus Hongo, Marasmius androsaceus (L.) Fr., Marasmius delectans Morgan, Marasmius pellucidus Berk. & Broome, Marasmius corbariensis (Roum.) Sacc., Marasmius rotula (Scop.) Fr., Marasmius fulvoferrugineus Gilliam, Marasmius sp.1, Marasmius sp.2, Marasmius sp.3, Marasmius sp.4, Marasmius sp.5, Marasmius sp.6, Tetrapyrgos nigripes (Fr.) E. Horak, Tetrapyrgos sp1, Tetrapyrgos sp.2. Trogia infundibuliformis Berk. & Broome, Vanromburghia silvetris Holterm, Gerronema sp. |
22 |
|
3 |
Mycenaceae |
Mycena stylobates (Pers.) P Kumm., Mycena alba (Bres.) Kühner, Mycena sp.1, Poromycena sp., Hydropus sp. |
5 |
|
4 |
Pseudoclitocybaceae |
Pseudoclitocybe cyathiformis (Bull.) Singer. |
1 |
|
5 |
Lyophyllaceae |
Termitomyces eurhizus (Berk.) R. Heim, Termitomyces clypeatus R. Heim, Termitomyces sp.1. |
3 |
|
6 |
Hygrophoraceae |
Hygrocybe coccineocrenata (PD Orton) MM Moser, Hygrocybe conica var. chloroides (Malençon) Bon, Hygrocybe psittacinavar. Perplexa (AH Sm & Hesler) Boertm, Hygrocybe ovina (Bull.) Kühner, Hygrocybe constrictospora Arnolds, Hygrocybe sp.1, Hygrocybe sp.2, Hygrocybe sp.3, Hygrocybe sp.4, Hygrocybe sp.5, Hygrocybe sp.6, Hygrocybe sp.7, Hygrophorus sp.1, Hygrophorus sp.1. Camarophyllus sp. |
15 |
|
7 |
Sarcoscyphaceae |
Cookeina sulcipes (Berk.) Kuntze, Cookeina tricholoma (Mont.) Kuntze, Cookeina sp.1, Phillipsia subpurpurea Berk. & Broome, Phillipsia domingensis (Berk.) Berk. ex Denison |
5 |
|
8 |
Agaricaceae |
Agaricus subrutilescens (Kauffman) Hotson & D.E. Stuntz, Agaricus pracelaresquamosus Freeman, Agaricus sp.1, Coprinus plicatilis (Curtis.) Fr., Coprinus cothurnatus Godey, Coprinus lagopus (Fr.) Fr., Coprinus cinereus (Schaeff.) Gray, Cyathus striatus Wiild., Cyathus pallidus Berk. & M.A.Curtis, Clarkeinda kuntze, Hymenagaricus alphitochrous (Berk.& Broome) Heinem, Leucocoprinus fragilissimus (Ravenel ex Berk & M.A Curtis) Pat. |
12 |
|
9 |
Psathyrellaceae |
Coprinellus disseminatus (Press.) J.E Lange |
1 |
|
10 |
Polyporaceae |
Hexagonia apiaria (Pers.) Fr., Microporus xanthopus (Fr.) Kuntze, Microporus sp, Picnoporus sanguineus (L.) Murrill, Picnoporus cinnabarinus (Jacq.) P. Karst., Trametes hirsute (Wulfen) Pilát, Trametes gibbosa (Pers.) Fr., Lenzites elegans (Spreng.) Pat., Trametes versicolor (L.) Lloyd, Lentinus ciliatus (Fr.) Zmitr, Lentinus bertieri (Fr.) Fr., Polyporus squamosus (Huds.) Fr., Laetiporus sulphureus (Bull.) Murrill. |
13 |
|
11 |
Russulaceae |
Russula atropurpurea (Krombh.) Britzelm., Russula cyanoxantha (Schaeff.) Fr., Russula aurata Fr., Russula sp.1, Russula sp.2, Russula sp.3, Lactifluus volemus (Fr.) Kuntze (1891) |
7 |
|
12 |
Amanitaceae |
Amanita phalloides (Vaill. Ex Fr.) Link, Amanita pantherina (DC.) Krombh., Amanita sp.1, Amanita sp.2 |
4 |
|
13 |
Xylariaceae |
Xylaria nigripes (Klotzsch) Cooke, Xylaria hypoxylon (L.) Grev., Xylaria sp.1, Xylaria sp.2, Xylaria sp.3, Xylaria sp.4, Xylaria sp.5 |
7 |
|
14 |
Hypoxylaceae |
Hypoxylon sp., Daldinia sp.. |
2 |
|
15 |
Auriculariaceae |
Auricularia polytricha (Mont.) Sacc., Auricularia delicate (Mont. ex Fr.) Henn., Auricularia auricula-judae (Bull.) Quél, Auricularia sp.1, Auricularia sp.2. Exidia recisa (Ditmar) Fr. |
6 |
|
16 |
Fomitopsidaceae |
Daedalea quercina (L.) Pers., Daedaleopsis confragosa (Bolton) J. Schröt, Daedaleopsis sp.1, Daedaleopsis sp.2 |
4 |
|
17 |
Strophariaceae |
Agrocybe retigera (Speg.) Singer, Agrocybe parasitica G. Stev, Agrocybe pediades (Fr.) Fayod |
3 |
|
18 |
Hymenochaetaceae |
Phellinus gilvus var. Licnoides (Mont.) Teng, Hymenochaete tabacina (Sowerby) Spirin, Miettinen & K.H. Larss, Hymenochaete rubiginosa (Dicks.) Lév. |
3 |
|
19 |
Geastraceae |
Geastrum mirabile Mont., Geastrum rufescens Pers. |
2 |
|
20 |
Pleurotaceae |
Pleurotus ostretaus P (Jack.) Kumm, Pleurotus sp. |
2 |
|
21 |
Tremellaceae |
Tremella fuciformis Berk., Tremella mesenterica Retz. |
2 |
|
22 |
Bolbitiaceae |
Panaeolus cyanescenssac. 1887, Conocybe tenera (Schaeff.) Kühner 1935 |
2 |
|
23 |
Schizophyllaceae |
Schizophyllum commune Fr., Schizophyllum sp.1 |
2 |
|
24 |
Hymenogastraceae |
Galerina sp., Gymnopilus sp. Psilocybe coprophila (Bull.) P. Kumm., |
3 |
|
25 |
Phalaceae |
Phallus sp.1, Phallus sp.2 |
2 |
|
26 |
Entolomataceae |
Entoloma nitidum Quėl. |
1 |
|
27 |
Gomphidiaceae |
Gomphidius roseus (Fr.) Oudem |
1 |
|
28 |
Holothiaceae |
Bisporella citrine (Batsch) Korf & SE Carp. |
1 |
|
29 |
Hydnaceae |
Clavulina sp.1, Clavulina sp.2 |
2 |
|
30 |
Corticiaceae |
Dendrothele sp. |
1 |
|
31 |
Stereaceae |
Stereum ostrea (Blume & T. Nees) Fr. |
1 |
|
32 |
Crepidotaceae |
Crepidotus uber (Berk. & MA Curtis) Sacc. |
1 |
|
33 |
Inocybaceae |
Inocybe sp. |
1 |
Figure 2. Distribution of macro mushrooms based on family differences
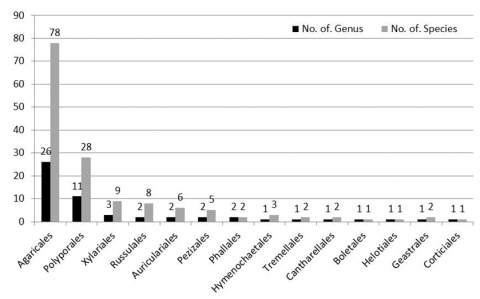
Furthermore, the order with the highest number of macro fungi is the order Agaricales as many as 78 species, then followed by the order Polyporales (28 species), Xylariales (9), Russulales (8), Auriculariales (6), (Pezizales 5), Hymenochaetales (3), Phallales, Tremellales, Cantharellales and Geastrales each have 2 species and Boletales, Helotiales and Corticiales also have 1 species (Figure 3). The most dominant order Agaricales was 52.7% (16 families, 26 genera and 78 species); order Polyporales 18.9% (3 families, 11 genera and 28 species); Xylariales 6.08% (2 families, 3 genera and 9 species); Russulales 5.4% (2 families, 2 genera and 8 species); Auriculariales 4.04% (1 family, 2 genera and 6 species); Pezizales 3.37% (1 family, 2 genera and 5 species); Hymenochaetales 2.02% (1 family, 1 genus and 3 species); Phallales 1.35% (1 family, 2 genera and 2 species); Tremellales 1.35% (1 family, 1 genus and 2 species); Cantharellales 1.35% (1 family, 1 genus and 2 species); Geastrales 1.35% (1 family, 1 genus and 2 species); Boletales 0.67% (1 family, 1 genus and 1 species); Helotiales 0.67% (1 family, 1 genus and 1 family); Corticiales 0.67% (1 family, 1 genus and 1 species) (Figure 4).
The results of this study are similar to those of Tibuhwa [57], where they found the dominant number of macro fungi of the order Agaricales and Polyprales in three types of land use in Tanzania. This is related to the fact that most types of macro fungi from members of the two orders are saprophytes, which have the ability to utilize available substrates and are able to degrade recalcitrant organic substrates [58, 59].
Figure 3. Distribution of macro mushrooms based on different orders
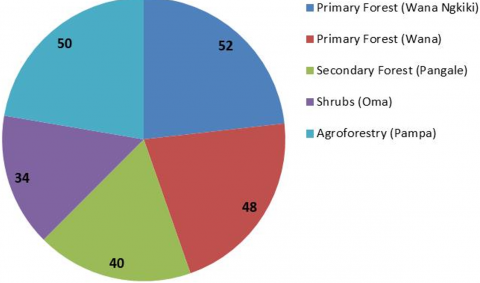
Diversity and species richness of macro mushrooms were found the most in primary forest (Wana Ngkiki and Wana) with 52 and 48 species respectively, followed by agroforestry land (Pampa) with 50 species, and decreased in secondary forest (Pangale) and shrubs (Oma) as many as 40 and 34 species, respectively (Figure 5). This may be due to the reduced number of plant species and the influence of human activities in the secondary forest (Pangale) and scrub (Oma).
Figure 4. Percentage of macro mushrooms based on different orders
Figure 5. Distribution of the number of types of macro fungi in different land use types
Andrew et al. [60] stated that succession involving changes in the composition of the fungal community is often associated with changes in the quality of the substrate. The number of macro fungi found in this study was 148 species, which is more than the study by Santos-Silva et al. [12], who found as many as 123 types of macro fungi (69 saprotrophic, 1 parasite and 53 mycorrhizal) in the Montado ecosystem, Southern Portugal or the results of research by Nacua et al. [61] which showed a smaller number of macro fungi, namely 21 macro fungal taxa on Mt. Makiling Forest Reserve (MMFR), Laguna, Philippines. Similar to the results of research by Ache et al. [62] which found the number of types of macro mushrooms found in the Kilum-Ijim Protection Forest.
Meanwhile, elsewhere, Lo´pez-Quintero et al. [63] reported a higher number of macro fungi, namely as many as 403 morphospecies of macro mushrooms belonging to 129 genera and 48 families of macro fungi found in the Colombian Amazon Forest. The results of research by Njuguini et al. [64] also showed a higher total number of macro fungi, namely 224 species divided into 28 families and 76 genera on several types of land use in Kereita Forest Kikuyu Escarpment, Kenya. Furthermore, Li et al. [65] found a larger number of macro-fungus species, namely 1360 species, which belong to 216 genera and 79 families in The Greater Mekong Sub region (GMS). They also found that natural forest had a higher diversity of macro-fungus species than plantation forest. The difference in the number of types of macro fungi in each type of land use is also caused by differences in the composition of the plant community and management methods. This is confirmed by Ferrer & Gilbert [66] that the species richness of macro fungi in an ecosystem has a strong relationship with tree species richness. Diversity and high plant density will result in a higher substrate diversity for macro fungi, which in turn causes a high diversity of macro fungi species because plants act as habitats and energy sources for most of these macro fungi [67, 68]. According to Gradstein et al. [69] that there are differences in the number of plant species that exist in each type of land use in Lore Lindu National Park Indonesia consists of 55 species in primary forest (Wana Ngkiki), 58 species in primary forest (Wana), 48 species in secondary forest (Pangale) and 19 species in agroforestry land.
However, other studies reveal that agroforestry systems that are managed with a diversified system are able to maintain biodiversity. In this study, what is interesting is that the agroforestry system (Pampa) has a high number of macro-fungus species compared to secondary forest (Pangale) and shrubs (Oma). This may be caused by the large number of leaves and tree wood that has died and then fallen to the forest floor, where during the study a lot of wood left over from tree cutting by farmers in processing their gardens was found on the forest floor. This is confirmed by Harley & Smith [70] who stated that changes in forest composition due to disturbance or timber harvesting will result in changes in the abundance and diversity of sporocarps, especially ectomycorrhizal fungi that depend on carbohydrates produced by their hosts. Furthermore, the large number of macro-fungus species in agroforestry areas is due to the large amount of waste wood bearings (logs) on the forest floor, which is a suitable substrate for many types of macro-fungus.
Many previous studies clearly document that in general the conversion of natural forest to other land use systems has a negative impact on diversity and species richness. Likewise in this study, the number of macro-mushroom species that were most commonly found in primary forest (Wana Ngkiki) was followed by primary forest (Wana), Pampa (agroforestry), secondary forest (Pangale) and shrubs (Oma) respectively.
It is very clear that the diversity of macro fungi is one indicator of ecosystem quality [71]. Angelini et al. [72] and Buée et al. [73] explained that the existence of different macro-fungal communities was associated with the dominant tree species in the forest. The diversity of tree species greatly affects the microclimate conditions in a place, which in turn affects the diversity and distribution of macro-mushroom species. Many studies report that the diversity and distribution of macro fungi in other regions found to be influenced by microclimatic conditions such as temperature, relative humidity, light, depth of decomposed organic matter (duff), humus, differences in sampling method, stand composition and soil conditions [12, 40, 74-81].
The index of diversity of macro-fungus species in several types of land use is presented in Table 3 and an example of the types of macro mushrooms found is presented in Figure 6.
Figure 6. Types of macro fungi found in several types of land use in Lore Lindu National Park
(a=Schizophyllum commune, b=Cookeina sp, c=Mycena sp, d=Microporus xanthopus, e=Clavulina sp., f=Phallus sp, g=Coprinellus disseminates, h=Geastrum mirabile, i=Boletus sp, j=Xylaria sp, k=Gymnopilus sp, l=Auricularia sp, m=Ganoderma sp, n=Rusulla sp, o=Marasmius sp, p=Stereum sp, q=Phillipsia sp, r=Daldinia sp, s=Tremella sp. t=Trametes vesicolor)
The Shannon and Simpson species diversity index in primary forest (Wana Ngkiki) with values of 1.70 and 0.98, respectively, was higher than in other land use types. There are 3 criteria for the value of the species diversity index, namely, if H' < 1 means low diversity, if H' = 1-3 means moderate diversity, if H` > 3 means high diversity. Based on these criteria, the diversity index of macro mushrooms in the five types of land use above is between 1.36-1.70 with a medium category (1-3), Due to the uneven distribution of macro fungi, there are certain types that can be found in very large numbers but there are certain species that are only found in very few. The higher the value of species diversity in an area, the more stable the community in the area is. High diversity indicates that a community has high complexity due to high species interactions. Furthermore, the uniformity index will reach its maximum value if the abundance of individuals per species is evenly distributed. The uniformity index value ranges from 0 to 1, the greater the value, the more uniform the number of individuals obtained. The smaller the uniformity index value, the smaller the species or genus uniformity in the community.
Table 3. Diversity index of macro fungi in several land use types around Lore Lindu National Park
|
Index/Value |
PFWK |
PFW |
SFP |
SO |
APP |
|
Simpson diversity index (1-D) Shannon diversity index (H') Evenness (EH) |
0.97 1.70 0.98 |
0.97 1.64 0.98 |
0.96 1.52 0.95 |
0.96 1.36 0.89 |
0.97 1.68 0.85 |
PFWK: Primary Forest (Wana Ngkiki); PFW: Primary Forest (Wana); SFP: Secondary Forest (Pangale); SO: Shrubs (Oma); APP: Agroforestry (Pampa)
According to Li et al. [82] and Priyamvada et al. [83] that the number of macro-fungus species will be more recorded in various regions if the environment is optimal depending on the season, temperature and amount of rainfall. The value of the diversity index of macro mushrooms obtained in the study may be higher if the research is carried out in optimal environmental conditions for the growth of macro mushrooms, therefore it is necessary to do this. exploration at different times and seasons each year.
This study also found several types of macro mushrooms that have the potential as food and medicine used by the Toro indigenous people who are the Kaili tribe, and are presented in the following Table 4.
Table 4. Types of macro mushrooms used by indigenous peoples in Toro village and its surrounding as a source of food and medicine
|
No. |
Local name |
Scientific name |
Used parts |
|
1. |
Tambata |
Pleurotus ostreatus |
The cap and stem of the fruiting body are used as food and medicine |
|
2. |
Tambata Talinga |
Auricularia sp |
The cap and stem of the fruiting body are used as food and medicine |
|
3. |
Tambata Talinga/ Lungo |
Auricularia auricula-judae |
The cap and stem of the fruiting body are used as food and medicine |
|
4. |
Tanggidi/ Tanggojo |
Schizophyllum commune |
The cap and stem of the fruiting body are used as food and medicine |
|
5. |
Rava |
Termytomyces sp |
The hood and stalk of the fruiting body as food |
|
6. |
Tambata |
Lentinus sp |
The hood and stalk of the fruiting body as food |
|
7. |
Tambata |
Agaricus subrutilescens |
The hood and stalk of the fruiting body as food |
|
8. |
Tambata |
Boletus sp |
The hood and stalk of the fruiting body as food |
|
9. |
Tambata goli |
Daldinia sp |
Fruit body as medicine |
|
10. |
Tambata |
Ganoderma lucidum |
Fruiting body as medicine |
As many as 10 types of macro mushrooms were found which were used as food and medicine by the indigenous people in Toro village. The number is less when compared to the study by De Leon et al. [84] which reported that there were 14 types of macro mushrooms used by the Aeta community in Central Luzon, Philippines. The processing of the macro mushroom fruiting bodies as food is carried out by local indigenous people by frying, sauteing and boiling with a mixture of other ingredients such as vegetables, fish, meat and spices so that they taste good for consumption. Among all these types of mushrooms, Schizophyllum commune (Tanggidi/Tanggojo) is the type of mushroom that is most often consumed by local people because apart from its delicious taste, it is also the easiest to grow and find. Some indigenous people in Toro village use the macro mushrooms above for medicinal purposes, such as medicine for coughs, ulcers, skin diseases, stomachaches and cancer. They usually boil or grind the fruiting bodies of the mushrooms and then the powder is mixed with coffee or the extract is used as a broth mixed with vegetables. There are also those whose fruit bodies are ground until smooth and then mixed with coconut oil and then applied directly to infected body parts such as boils and pimples. The use of macro mushrooms as food and medicine is almost the same as the method used by the indigenous Bugkalot people in Alfonso Castañeda, Nueva Vizcaya, Philippines [85, 86].
Macro mushrooms that are large in Kaili language are called Tambata, while the small ones are called Tanggidi, some call them Tanggojo (34). Naming also follows its shape, for example macro mushrooms shaped like ears are called Tambata Talinga (Talinga = Ears), those shaped like marbles are called Tambata goli (Goli = Males). Similar to the naming of macro mushrooms by Njouonkou et al. [87] in the Cameron West Region, it is also based on their shape, for example the shape of fruits such as melons and also based on the substrate where they grow, for example Tambata Tai Japi (In Kaili language Tai = Dirt, Japi- = Cow), and so on.
Our results confirm diverse macro mushrooms community in some land use types around Lore Lindu National Park, and need for their conservation. A total of 148 types of macro mushrooms were found in five types of land use. Conversion of primary forest to secondary forest, agroforestry and shrubs altered macro mushrooms species composition, but did not affect species diversity. The highest number of macro-mushroom species was found in primary forest (Wana Ngkiki) with 52 species, followed by primary forest (Wana) with 48 species, agroforestry land (Pampa) with 50 species, secondary forest (Pangale) with 40 species and the least found in shrubs (oma) as many as 34 species. Several types of macro mushrooms were found to have potential as food and medicine, which are often used by local indigenous peoples, namely Pleurotus sp, Auricularia sp, Auricularia auricula-judae, Termytomyces sp, Lentinus sp, Schizophyllum commune, Agaricus subrutilescens, Boletus sp, Daldinia sp and Ganoderma lucidum. The number of types of macro mushrooms found in this area is certainly not a final investigation. Therefore, further expeditions to explore the diversity of macro fungi should be carried out in all parts of the park and in different seasons every year in order to obtain more complete data of macro fungi in the garden and the possibility to obtain new species or records in this park.
This research was funded by the Directorate of Community Service and Research, Ministry of Research, Technology and Higher Education, Republic of Indonesia, through the Basic Research scheme with contract number 023.04.1.673453/2015. The author also expresses his deepest gratitude to the community and Toro village government for their participation and support in this research, especially for the research permit and support in providing lodging and logistics, especially to Mr. Said for the assistance of the research team in the field. We also thank Azmul, Rustam and the Wana Cikal team, Faculty of Forestry, Tadulako University (Foiske Tamporo, Moh Fadhil, Viviyen, Berwin Toheke, Roy and Christian) for their assistance in conducting research in the field.
[1] UNESCO. (1995). Co-operative scientific study of East Asian biosphere reserves. United Nations Educational, Scientific and Cultural Organization, Jakarta.
[2] TNC/BTNLL. (2002). Lore Lindu National Park. Draft Management Plant 2002-2007. Direktorat Jenderal Perlindungan Hutan dan Konservasi Alam and The Nature Conservancy, Palu, Indonesia.
[3] Ramadhanil. (2006). Strucuture and composition of vegetation in six land use types in the Lore Lindu National Park, Central Sulawesi, Indonesia. Dissertation. Department of Biology. The Graduate School, Bogor Agricultural University.
[4] Muchane, M.N., Muchane, M., Mugoya, C., Masiga, C.W. (2012). Effect of land use system on arbuscular mycorrhiza fungi in Maasai Mara Ecosystem, Kenya. African Journal of Microbiology Research, 6(17): 3904-3916. https://doi.org/10.5897/AJMR12.155
[5] Das, K. (2010). Diversity and conservation of wild mushrooms in Sikkim with special reference to Barsey Rododendron Sanctuary. NeBio, 1(2): 1-13.
[6] Haryati, J.R., Azrianingsih, R. (2012). Ethno-Edible Mushroom of Pleurotus sp., Clytocybenebularis and Auricularia auricular in Ranupani Village, East Java. The Journal of Tropical Life Science, 2(2): 40-43.
[7] Hawksworth, D.L., Lücking, R. (2017). Fungal diversity revisited: 2.2 to 3.8 million species. In: Heitman J., Howlett B.J., Crous P.W., Stukenbrock E.H., James T.Y., Gow, N.A.R. Fungal Kingdom. AMS Press. Washington DC, 79-95. https://doi.org/10.1128/9781555819583.ch4
[8] Hawksworth, D.L. (2001). The magnitude of Fungal Diversity: The 1.5 million species estimate revisited. Mycological Research, 105: 1422-1432. https://doi.org/10.1017/S0953756201004725
[9] Mueller, G.M., Schimt, J.P., Leacock, P.R., Buyk, B., Cifuentes, J., Desjardin, D.E., Halling, R.E., Hjortstam, K., Iturruaga, T., Larsson, K.H., Lodge, D.J., May, T.W., Minter, D., Rajchenberg, M., Redhead, S.A., Ryvarden, L., Trappe, J.M., Watling, R., Wu, Q. (2007). Global diversity and distribution of macrofungi. Biodiversity and Conservation, 16: 37-48. https://doi.org/10.1007/s10531-006-9108-8
[10] Chang, S., Miles, G.P. (2004). Mushrooms: Cultivation, Nutritional Value, Medicinal Effects and Environmental Impact (p. 436). Boca Raton, FL: CRC Press. http://dx.doi.org/10.1201/9780203492086
[11] Rossman, A.Y. (1994). A strategy for an all-taxa inventory of fungal biodiversity. In: C.I Peng and C.H Chou (eds.). Biodiversity and Terrestrial Ecosystem. Institute of Botany, Academia Sinica, Taipei, Taiwan, 14: 169-194.
[12] Santos-Silva, C., Gonçalves, A., Louro, R. (2011). Canopy cover influence on macrofungal richness and sporocarp production in Montado ecosystems. Agroforest System, 82: 149-159. https://doi.org/10.1007/s10457-011-9374-7
[13] Cannon, P.F., Aguirre-Hudson, B., Aime, M.C., Ainsworth, A.M., Bidartondo, M.I., Gaya, E., Hawksworth, D., Kirk, P., Leitch, I.J., Lucking, R. (2018). Chapter 1: Definition and diversity. 4-11. In: State of the World's Fungi 2018. Royal Botanic Gardens Kew.
[14] Roosheroe, I.G., Sjamsuridzal, W., Oetrai, A. (2006). Mikologi Dasar dan Terapan. Yayasan Pustaka Obor Jakarta, Indonesia.
[15] Corner, E.J.H. (1996). The agaric genera Marasmius, Chaetocalathus, Crinipellis, Heimiomyces, Resupinatus, Xerula and Xerulina in Malesia. Beiheft zur Nova Hedwigia, 111: 1-175
[16] Lee, S.S., Besl, H., Salmiah, U. (1995). Some of the fungi of the Halong River and surrounding areas, Temenggor Forest Reserve, Hulu Perak, Malaysia. Malaysian Nature Journal, 48: 147-155.
[17] Desjardin, D.E., Flegel, T.W., Boonpratuang, T. (2004). Basidiomycetes. In: EGB Jones, M Tanticharoen and KD Hyde (eds.). Thai Fungal Diversity, Biotech Publishing, Thailand, 37-49.
[18] Khastini, R.O., Wahyuni, I., Lista., S. (2019). Inventory and utilization of macrofungi species for food in Cikartawana inner Baduy Banten. Biodidaktika: Jurnal Biologi dan Pembelajarannya, 14(1): 7-13. https://doi.org/10.30870/biodidaktika.v14i1.4838
[19] Lu, H., Lou, H., Hu, J., Liu, Z., Chen, Q. (2020). Macrofungi: A review of cultivation strategies, bioactivity, and application of mushrooms. Comprehensive Reviews in Food Science and Food Safety, 19: 2333-2356. https://doi.org/10.1111/1541-4337.12602
[20] Putra, I.P. (2020). Record on macroscopic fungi at IPB university campus forest: Description and potential utilization. Indonesian Journal of Science and Education, 4(1): 1-11. http://dx.doi.org/10.31002/ijose.v4i1.2180
[21] Valverde, M.E., Hernández-Pérez, T., Paredes-López, O. (2015). Edible mushrooms: Improving human health and promoting quality life. International Journal of Microbiology, 1-14. https://doi.org/10.1155/2015/376387
[22] De la Parra, B., Armenteros, S., Cuesta, J., Olaizola, J., Santos, L., Monléon, V., Herrero, C. (2020). Sprouting suppression and mushroom production after inoculation of Juglans x intermedia stumps with edible fungi species. Annals of Silvicultural Research, 44(1): 30-40. https://doi.org/10.12899/asr-1936
[23] Putra, I.P. (2020). Allelopathic activity of some wild mushrooms in Indonesia. Fungal Territory, 3(1): 1-3. https://doi.org/10.36547/ft.2020.3.1.1-3
[24] United Nations Development Programme. (2012). Ngata Toro Community, Indonesia. Equator Initiative Case Study Series. New York, NY.
[25] Ariyanti, N.S., Bos, M.M., Kartawinata, K., Tjitrosoedirdjo, S.S., Guhardja, E., Gradstein, S.R. (2005). Bryophytes on tree trunks in natural forests, selectively loggeds forests and cacao agroforests in Central Sulawesi. Biological Conservation, 141: 2516-2527.
[26] Gradstein, S.R., Tan, B.C., King, C., Zhu, R.L., Drubert, C., Pitopang, R. (2005). Catalog of the bryopytes of Sulawesi, Indonesia. Journal of the Hattori Botanical Laboratory, 98: 213-257.
[27] Kessler, M., Kessler, P.J.A., Gradstein, S.R., Bach, K., Schmull, M., Pitopang, R. (2005). Tree diversity in primary forest and different land use systems in Central Sulawesi, Indonesia. Biodiversity and Conservation, 14: 547-560. https://doi.org/10.1007/s10531-004-3914-7
[28] Maryanto, I., Yani, M., Priyono, S.N., Wiantoro, A. (2011). Altitudinal distribution of fruit bats (Pteropodidae) in Lore Lindu National Park, Central Sulawesi, Indonesia. Hystrix Italian Journal of Mamalogy, 22(1): 167-177. https://doi.org/10.4404/Hystrix-22.1-4480
[29] Schulze, C.H., Waltert, M., Kessler, P.J.A., Pitopang, R., Shahabuddin, Veddeler, D., Muehlenberg, M., Gradstein, S.R., Leuschner, C., Steffan-Dewenter, I., Tscharntke, T. (2004). Biodiversity indicator taxa of tropical land- use systems: comparing plants, birds and insects. Ecological Applications, 14(5): 1321-1333. https://doi.org/10.1890/02-5409
[30] Wanger, T.C., Saro, A., Iskandar, D.T., Motzke, I., Brook, B.W., Sodhi, N.S., Clough, Y., Tscharntke, T. (2009). Conservation value of cacao agroforestry for amphibians and reptiles in South-East Asia: combining correlative models with follow- up field experiments. Journal of Applied Ecology, 46: 823-832. https://doi.org/10.1111/j.1365-2664.2009.01663.x
[31] Wanger, T.C., Iskandar, D.T., Brook, B.W., Sodhi, N.S., Clough, Y., Tscharntke, T. (2010). Effects of land-use change on community composition of tropical amphibians and reptiles in Sulawesi, Indonesia. Conservation Biology, 24(3): 795-802. https://doi.org/10.1111/j.1523-1739.2009.01434
[32] Wanger, T.C., Rauf, A., Schwarze, S. (2010). Pesticides and tropical biodiversity. Frontiers in Ecology and the Environment, 8(4): 178-179. https://doi.org/10.1890/10.WB.015
[33] Wielgoss, A., Tscharntke, T., Buchori, D., Fiala, B., Clough, Y. (2010). Temperature and a dominant dolichoderine ant species affect ant diversity in Indonesian cacao plantations. Agriculture, Ecosystems and Environment, 135: 253-259. https://doi.org/10.1016/j.agee.2009.10.003
[34] Yusran, Y., Erniwati, E., Wahyuni, D., Ramadhanil, R., Khumaidi, A. (2021). Diversity of macro fungus across three altitudinal ranges in Lore Lindu National Park, Central Sulawesi, Indonesia and their utilization by local residents. Biodiversitas, 22(1): 199-210. https://doi.org/10.13057/biodiv/d220126
[35] Ramadhanil, Tjitrosoedirdjo, S.S., Setiade, D. (2008). Structure and composition of understory plant assemblages of six land use types in the Lore Lindu National Park, Central Sulawesi, Indonesia. Bangladesh Journal of Plant Taxonomy, 15(1): 1-12. https://doi.org/10.3329/bjpt.v15i1.911
[36] Falk, U., Ibrom, A., Oltchev, A., Kreilein, H., June, T., Rauf, A., Merklein, J., Gravenhorst, G. (2005). Energy and water fluxes above a cacao agroforestry system in Central Sulawesi Indonesia, indicating the effects of land use change on local climate. Meteorologische Zeitschrift, 14(2): 219-225. https://doi.org/10.1127/0941-2948/2005/0025
[37] Gerold, G., Fremerey, M., Guhardja, E. (2004). Land use, nature conservation, and the stability of rainforest margins in Southeast Asia. Springer-Verlag Berlin Heildelberg New York, London, Paris and Tokyo.
[38] Whitten, T., Mustafa, M., Henderson, G.S. (1987). The Ecology of Sulawesi. Gadjah Mada University Press, Yogyakarta.
[39] Yuliana, Sriyati, S., Sanjaya, Y. (2017). Local wisdom of Ngata Toro community in utilizing forest resources as a learning source of biology. AIP Conference Proceedings 1868, 100007. https://doi.org/10.1063/1.4995217
[40] Engola, A.P.O., Eilu, G., Kabasa, J.D., Kisovi, L., Munishi, P.K.T, Olila, D. (2007). Ecology of edible indigenous mushrooms of the Lake Victoria Basin, Uganda. Research Journal of Biological Sciences, 2(1): 62-68.
[41] Brown, N., Bhagwat S., Watkinson S. (2006). Macrofungal diversity in fragmented and disturbed forests of the Western Ghats of India. Journal of Applied Ecology, 43: 11-17. https://doi.org/10.1111/j.1365-2664.2005.01107.x
[42] Osemwegie, O.O., Okhuoya, J.A. (2011). Diversity and abundance of macrofungi in rubber agroforests in southwestern Nigeria. Nordic Journal of Botany, 29: 119-128. https://doi.org/10.1111/j.1756-1051.20100.00717.x
[43] Barnett, H.L., Hunter, B.B. (1998). Illustrated genera of imperfect fungi. The American phytopathological society. US Department of Agriculture, Agricultural Research Service, Washington State University, Pullman. APS Press. USA. St. Paul, Minnesota USA. 218p.
[44] Desjardin, D.E., Retnowati, A., Horak, E. (2000). Agaricales of Indonesia 2. A Preliminary monograph of Marasmius from Java and Bali. Sydovia, 52(2): 92-193.
[45] Fassatiova, O. (1986). Molds and filamentous fungi in technical microbiology. Department of Cryptogamic Botany Charles University. Prague.
[46] Hemmes, D.E., Desjardin, D.E. (2002). Mushrooms of Hawai'I (An identification guide). Ten speed press. Berkeley, California. 212.
[47] Härkönen, M., Saarimäki, T., Mwasumbi, L. (1995). Edible mushrooms of Tanzania. Karstenia (Supplements), 35: 1-92. https://doi.org/10.29203/ka.1995.314
[48] Jordan, M. (2004). The Encyclopedia fungi of Britain and Europe. Frances Lincoln Ltd. London, 384.
[49] Kirk, P.M., Cannon, P.F., David, J.C., Stalpers, J.A. (2001). Ainsworth & Bisby's dictionary of the fungi: 9th Edition. CABI Publishing. Wallingford, UK.
[50] Largent, D.L. (1986). How to identify mushrooms into genus I: Macroscopic Features. Mad River Press Inc. California.
[51] Laessøe, T., Lincoff, G. (2002). Mushrooms. Smithsonian Handbooks. Dorling Kinderslay Inc. New York.
[52] Savonius, M. (1973). All Color Book of Mushrooms and Fungi. Octopus Books. London.
[53] Index Fungorum. (2015). http://www.indexFungorum.org/names, accessed on 22 February 2022.
[54] Margalef, R. (2008). Correspondence between the classic types of lakes and the structural and dynamic properties of their populations. Internationale Vereinigung fuer Theoretische und Angewandte Limnologie, 15: 169-170.
[55] Simpson, E. (1949). Measurement of Diversity. Nature. 163: 688. https://doi.org/10.1038/163688a0
[56] Pielou, E.C. (1996). The measurement of diversity in different gypes of biological collections. Journal of Theoretical Biology, 13: 131-144. https://doi.org/10.1016/0022-5193(66)90013-0
[57] Tibuhwa, D.D. (2011). Diversity of Macro Fungi at the University of Dar es Salaam Mlimani main campus in Tanzania. International Journal of Biodiversity and Conservation, 3(11): 540-550.
[58] Lynch, M.D.J., Thorn, R.G. (2006). Diversity of basidiomycetes in Michigan agricultural soils. Applied Environment Microbiology, 72(11): 7050-7056. https://doi.org/10.1128/AEM.00826-06
[59] Osemwegie, O.O., Eriyaremu, E.G., Abdulmalik, J. (2006). A survey of macrofungi in the Edo/Delta region of Nigeria, their morphology and uses. Global Journal of Pure and Applied Sciences, 12(2): 149-157. https://doi.org/10.4314/gjpas.v12i2.16583
[60] Andrew, E.E., Kinge, T.R., Tabi, E.M., Thiobal, N., Mih, A.M. (2013). Diversity and distribution of macrofungi (mushrooms) in the Mount Cameron region. Journal of Ecology and the Natural Environment, 5(10): 318-334.
[61] Nacua, A.E., Pacis, H.J.M., Manalo, J.R., Soriano, C.J.M., Tosoc, N.R.N., Padirogao, R., Clemente, K.J.E., Deocaris, C.C. (2018). Macrofungal diversity in Mt. Makiling Forest Reserve, Laguna, Philippines: with floristic update on roadside samples in Makiling Botanic Gardens (MBG). Biodiversity, 19: 1579-1585. https://doi.org/10.13057/biodiv/d190451
[62] Ache, N.T., Kinge, T.R., Bechem, E.E.T., Ndam, L.M., Mih, A.M. (2019). Mushroom species richness, distribution and substrate specificity in the Kilum-Ijim forest reserve of Cameroon. Journal of Applied Biosciences, 133: 13592-13617. https://dx.doi.org/10.4314/jab.v133i1.11
[63] López-Quintero, C.A., Straatsma, G., Franco-Molano, A.E., Boekhout, T. (2012). Macrofungal diversity in Colombian Amazon forests varies with regions and regimes of disturbance. Biodiversty and Conservation, 21: 2221-2243. https://doi.org/10.1007/s10531-012-0280-8
[64] Njuguini, S.K.M., Muchane, M.N., Wachira, P., Okoth, S., Muchane, M., Saado, H. (2018). Effects of land use on the diversity of Macrofungi in Kereita Forest Kikuyu Escarpment, Kenya. Current Research in Environmental & Applied Mycology, 8(2): 254-281. https://doi.org/10.5943/cream/8/2/10
[65] Li, H., Guo, J., Karunarathna, S.C., Ye, L., Xu, J., Hyde, K.D., Mortimer, P.E. (2018). Native forests have a higher diversity of Macrofungi than comparable plantation forests in the greater Mekong subregion. Forests, 9(7): 1-14. https://doi.org/10.3390/f9070402
[66] Ferrer, A., Gilbert, G.S. (2003). Effect of tree host species on fungal community composition in a tropical rain forest in Panama. Diversity and Distribution, 9: 455-468. https://doi.org/10.1046/j.1472-4642.2003.00039.x
[67] Lodge, D.J., Ammirati, J.F., O'Dell, T.E., Mueller, G.M. (2004). Collecting and describing macrofungi. Biodiversity of fungi: inventory and monitoring methods. Amsterdam: Elsevier Academic Press, 2004: 128-158.
[68] Schmit, J.P., Mueller, G.M., Leacock, P.R., Mata, J.L., Wu, Q., Huang, Y. (2005). Assessment of tree species richness as a surrogate for macrofungal species richness. Biological Conservation, 121: 99-110. https://doi.org/10.1016/j.biocon.2004.04.013
[69] Gradstein, S.R., Kessler, M., Pitopang, R. (2007). Tree species diversity relative to human land uses this tropical rain forest margins in Central Sulawesi. In: Tscharntke T, Leuschner C, Zeller M, Guhardja E, Bidin A (eds), The stability of tropical rainforest margins, linking ecological, economic and social constraints of land use and conservation, Springer Verlag Berlin, 321-334. https://doi.org/10.1007/978-3-540-30290-2_16
[70] Harley, J.L., Smith, S.E. (1983). Mycorrhizal Symbiosis. Academic Press, London, 483.
[71] Stamets, P. (2000). The role of mushrooms in nature, culturing mushroom mycelium on agar media. In: Growing Gourmet and medicinal mushrooms. Ten Speed Press, Berkeley. CA.
[72] Angelini, P., Giancarlo, B., Andrea, A., Emma, B., Roberto, V. (2015). Diversity and ecological distribution of macrofungi in a site of community importance of Umbria (Central Italy). The Open Ecology Journal, 8: 1-8.
[73] Buée, M., Maurice, J.P., Zeller, B., Andrianarisoa, S., Ranger, J., Courtecuisse, R., Marçaise, B., Le Tacon, F. (2011). Influence of tree species on richness and diversity of epigeous fungal communities in a French temperate forest stands. Fungal Ecology, 4: 22-31.
[74] Alem, D., Dejene, T., Oria-de-Rueda, J.A., Martín-Pinto, P. (2021). Survey of macrofungal diversity and analysis of edaphic factors influencing the fungal community of church forests in dry Afromontane areas of Northern Ethiopia. Forest Ecology and Management, 496: 1-16. https://doi.org/10.1016/j.foreco.2021.119391
[75] Baptista, P., Martins, A., Tavares, R.M., Lino-Neto, T. (2010). Diversity and fruiting pattern of macrofungi associated with chestnuts (Castanea sativa) in the Trásos Montes region (Northeast Portugal). Fungal Ecology, 3: 9-19. https://doi.org/10.1016/j.funeco.2009.06.002
[76] Bergemann, S.E., Largent, L.D. (2000). The site-specific variables that correlates with the distribution of the Pacific Golden Chanterelle (Cantharellus formossus). Forest Ecology and Management, 130: 99-107. https://doi.org/10.1016/S0378-1127(99)00177-2
[77] Conn, C., Dighton, J. (2000). Litter quality influences on decomposition, ectomycorrhizal community structure and mycorrhizal root surface acid phosphatase activity. Soil Biology & Biochemistry, 32: 489-496. https://doi.org/10.1016/S0038-0717(99)00178-9
[78] Ferris, R., Peace, A.J., Newton, A.C. (2000). Macrofungal communities of lowland Scots pine (Pinus sylvestris L.) and Norway spruce (Picea abies (L.) Karsten.) plantations in England: relationships with site factors and stand structure. Forest Ecology and Management, 131: 255-267.
[79] Packham, J.M., May, T.W., Brown, M.J., Wardlaw, T.J., Mills, A.K. (2002). Macrofungal diversity and community ecology in mature and regrowth wet eucalpt forest in Tasmania: A multivariate study. Austral Ecology, 27: 149-161. https://doi.org/10.1046/j.1442-9993.2002.01167.x
[80] Pushpa, H., Purushothama, K.B. (2012). Biodiversity of mushrooms in and around Bangalore (Karnataka), India. American-Eurasian Journal of Agricultural and Environmental Sciences, 12(6): 750-759. https://doi.org/10.5829/idosi.aejaes.2012.12.06.56401
[81] Suggitt, A.J., Gillingham, P.K., Hill, J.K., Huntley, B., Kunin, W.E., Roy, D.B., Thomas, C.D. (2011). Habitat microclimates drive fine-scale variation in extreme temperatures. Oikos, 120(1): 1-8. https://doi.org/10.1111/j.1600-0706.2010.18270.x
[82] Li, H., Guo, J., Goldberg, S.D., Sreekar, R., Ye, L., Luo, X., Sysouphanthong, P., Xu, J., Hyde, K.D., Mortimer, P.E. (2018). Fruiting patterns of macrofungi in tropical and temperate land use types in Yunnan Province, China. Acta Oecologica, 91: 7-15. https://doi.org/10.1016/j.actao.2018.05.008
[83] Priyamvada, H., Akila, M., Singh, R.K., Ravikrishna, R., Verma, R.S., Philip, L., Marathe, R.R., Sahu, L.K., Sudheer, K.P., Gunthe, S.S. (2017). Terrestrial macrofungal diversity from the tropical dry evergreen biome of Southern India and its potential role in aerobiology. Plos One, 12(1): 1-21. https://doi.org/10.1371/journal.pone.0169333
[84] De Leon, A.M., Reyes, R.G., dela Cruz, T.E.E. (2012). An ethnomycological survey of macrofungi utilized by Aeta communities in Central Luzon, Philippines. Mycosphere, 3(2): 251-259.
[85] Torres, M.L.S., Ontengco D.C., Tadiosa E.R., Reyes R.G. (2020). Ethnomycological studies on the bugkalot indigenous community in Alfonso Castañeda, Nueva Vizcaya, Philippines. International Journal of Pharmaceutical Research and Allied Science, 9(4): 43-54.
[86] Arifah, S.M., Budiastuti, M.T.S., Dewi, W.S., Supriyadi. (2021). Vermicompost formulation based on soybean husk and cow manure on shallots. International Journal of Design & Nature and Ecodynamics, 16(3): 327-333. https://doi.org/10.18280/ijdne.160312
[87] Njouonkou, A.L., Crop, E.D., Mbenmoun, A.M., Kinge, T.R., Biyé, E.H., Verbeken, A. (2016). Diversity of edible and medicinal mushrooms used in the noun division of the west region of Cameroon. International Journal of Medicinal Mushrooms, 18(5): 387-396. https://doi.org/10.1615/IntJMedMushrooms.v18.i5.20