Quality of the Multi-scale Interphase of Hemp Stems: Retting Effect
Qualité de l’Interphase Multi-échelle des Tiges de Chanvre : Effet du Rouissage

Samuel Réquilé* | Antoine Le Duigou | Alain Bourmaud | Christophe Baley
OPEN ACCESS
The control of hemp plant harvesting, in particular the retting process, represents the major step for the development of high-performance hemp biocomposites. In this study, the consequences of hemp retting on the ease of fibre extraction and tensile properties of elementary hemp fibres is explored. Stem peeling experiments are combined with scanning electron microscopy (SEM) to investigate the influence of the retting degree in plants. After studying the evolution of fracture energy at the fibre/woody core interphase within the stem, this work investigates interphase fracture mechanisms during peeling experiments which aims to understand the evolution of fibre bundle cohesion during retting. A drastic drop of fracture energy reveals the impact of retting on the ease of peeling the outer tissue of the stem (containing fibre bundles). The positive impact of retting on various fibre defects combined together leads to a 33 % increase in the tangent modulus between non-retted and retted fibres, highlighting the importance of understanding the retting process to open up to new markets for biocomposites.
RÉSUMÉ :
Le contrôle de la récolte de la plante de chanvre, en particulier le rouissage, représente l'étape majeure pour le développement de biocomposites renforcés par des fibres de chanvre à hautes performances. Dans cette étude, les conséquences du rouissage du chanvre sur la facilité d’extraction des fibres ainsi que leurs propriétés mécaniques longitudinales sont explorées. Des essais de pelage de tiges sont combinés avec des images de microscopie électronique à balayage (MEB) pour étudier l'influence du degré de rouissage dans les plantes. Après avoir étudié l’évolution de l’énergie de rupture au niveau de l’interphase fibre/xylème au sein de la tige, ce travail s’intéresse aux mécanismes de rupture de cette interphase lors du pelage afin de comprendre l’évolution de la cohésion des faisceaux de fibres lors du rouissage. Une chute drastique de l'énergie de rupture révèle l'impact du rouissage sur la dégradation de l’interphase et donc la facilité à peler le tissu externe de la tige (contenant les faisceaux de fibre). Ceci conduit à une augmentation de 33 % du module tangent entre les fibres non rouies et les fibres rouies, soulignant l’importance de la compréhension du processus de rouissage pour ouvrir de nouveaux marchés aux biocomposites.
biocomposites, hemp, fiber, interface, retting, peeling test
Mots-clés :
biocomposite, chanvre, fibre, interface, rouissage, test de pelage
L'utilisation de fibres végétales est une réelle opportunité pour satisfaire une demande croissante de matériaux éco-conçus performants. Bien que principalement destinées à l'industrie papetière, les fibres de chanvre possèdent des caractéristiques mécaniques intrinsèques qui favorisent leur valorisation potentielle dans de nouvelles applications industrielles techniques et à forte valeur ajoutée, notamment les non-tissés pour pièces thermo-compressées, les matériaux composites injectés et les textiles [1-4]. L'accès à ces principaux marchés implique cependant la production de matières premières de haute qualité répondant à des spécifications strictes. Ceci nécessite la transformation des pailles de chanvre avant le défibrage par rouissage des tiges, inspirée des cultures ciblées sur le textile telles que le lin. De nombreuses études ont déjà démontré l'impact de la transformation des tiges [5-7], et du rouissage sur la qualité des fibres en terme de composition biochimique, de comportement mécanique, de propriétés de surface et de cohésion des éléments fibreux [8-10]. Certains auteurs ont souligné l’impact de l’étape d’extraction effectuée sur des tiges vertes, qui affectent les propriétés mécaniques des fibres obtenues, engendrant divers défauts dans leur microstructure [11-14]. Pour les limiter, la solution consiste à faciliter le défibrage en adaptant les conditions de rouissage en fonction de l'application ciblée.
Abandonné au milieu du XXème siècle en raison de la pollution de l'eau (eutrophisation) par les produits de fermentation des tiges, le rouissage à l'eau a été remplacé par le rouissage au champ pour la culture des plantes. L’étape de rouissage pourrait entraîner une séparation préalable des fibres du reste de la tige et au sein des faisceaux en dégradant les constituants pectiques présents dans les lamelles mitoyennes [15-17]. Actuellement, les producteurs de chanvre déterminent de façon très empirique le degré de rouissage obtenu en utilisant des propriétés organoleptiques (essais mécaniques manuels, couleur, dureté des tiges). Ces paramètres ne reflètent que très partiellement la qualité des fibres et sont perfectibles. Ainsi, il est très difficile pour les producteurs de chanvre de fournir des pailles dont le niveau de rouissage est contrôlé pour répondre aux spécifications des applications visées. Compte tenu de l’importance de l’impact du rouissage sur la qualité des fibres de chanvre et des exigences des industries utilisatrices de fibres, il est essentiel de définir des indicateurs permettant de mesurer le degré de qualité de la matière première.
L'objectif principal de cette étude est d'utiliser des essais de pelage de tige pour étudier l'effet du rouissage sur la cohésion tissulaire des tiges de chanvre et son impact sur les propriétés longitudinales des fibres élémentaires. La littérature existante mentionne des méthodes permettant d'évaluer le degré de rouissage en étudiant plusieurs facteurs : composition chimique, morphologie, colonisation bactérienne / fongique [18-21]. Cependant, ces méthodes ne permettent pas de quantifier de manière mécanique la qualité de l’interphase, ou capacité de décohésion des tiges, comme cela est fait dans cet article. Tout d'abord, les conditions de pelage et la teneur en humidité des échantillons sont déterminées avec précision pour assurer une comparaison valable entre des échantillons similaires. Des études antérieures [22-25], ont montré l'influence de l'état de l'échantillon (teneur en humidité) et des conditions de pelage telles que la température, la vitesse et l’angle de pelage sur le comportement de rupture et la force de pelage des adhésifs. Après avoir décrit les phénomènes de pelage dans les tiges présentant différents degrés de rouissage, et combinés à des observations microscopiques des surfaces pelées, une analyse de l'énergie de rupture à l'interphase fibre/bois est présentée. Enfin, l’impact du degré de rouissage sur les performances mécaniques des fibres élémentaires est exploré.
2.1 Materials
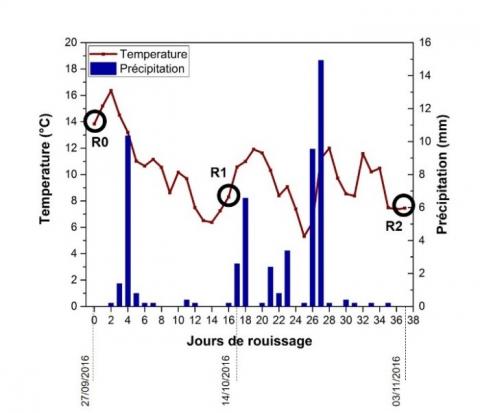
Des tiges de chanvre (Cannabis sativa L., cultivar Fedora 17) ont été cultivées à La Chanvrière (Bar-sur-Aube, France). Les tiges ont été récoltées mécaniquement fin août 2016 après récolte des graines. Les tiges ont ensuite été laissées au champ pendant 37 jours pour l’étape de rouissage. 10 tiges ont été prélevées au hasard avant le rouissage (échantillon R0), après 17 jours de rouissage (R1) et après 37 jours (R2) pour cette étude, puis séchées naturellement à l'air (cf. Figure 1). Les tiges ont été maintenues dans un environnement contrôlé (T = 23 °C et HR = 50 %) avant tout essai. Les fibres ont été extraites à l'aide d'un procédé mécanique industriel qui consiste à briser et à teiller les pailles.
2.2 Teneur en eau des tiges
Les tiges ont été maintenues dans un environnement contrôlé (T=23 °C et RH=50 %) pendant 48 heures pour stabilisation de leur masse et teneur en eau. Les tiges ont ensuite été séchées à l'étuve à une température de 105 °C (température communément utilisée pour l’élimination d’eau libre et d’eau liée [26] sous pression atmosphérique jusqu'à ce que l’échantillon atteigne une masse constante. Des échantillons de tiges ont été périodiquement retirés de l’étuve pour être pesés. Le pourcentage de perte d'eau au sein des tiges, notés Mt, a été calculé à l'aide de la formule suivante (Eq. 1) :
$M_{t}=\frac{w_{0}-w_{t}}{w_{0}}$ (1)
où W0 et Wt sont la masse des tiges avant séchage et la masse du matériau sec, respectivement.
Tous les échantillons montrent une teneur en eau moyenne de 8,5 ± 0,3 %. Seule une légère variation de 0,5 % a été observée entre les échantillons présentant différents degrés de rouissage. Des études antérieures ont montré l'influence de la variation de la teneur en eau du bois sur la résistance à la rupture [27], atteignant une valeur maximale à une humidité relative de 82 %. La résistance à la rupture augmente lorsque le matériau sèche et devient moins ductile, atteignant un maximum de 16,8 % d’humidité. Booth et al. [28] ont également observé l'influence de la teneur en eau de la tige sur le travail nécessaire pour peler une tige en raison des modifications des propriétés mécaniques des parois cellulaires. Dans la présente étude, nous considérons que toutes les tiges ont des teneurs en eau similaires, ces dernières ayant subi les mêmes conditions de stockage (voir section 2.1).
Figure 1. Températures et précipitation moyennes quotidiennes pendant la période de rouissage des tiges de chanvre
2.3 Essais de pelage
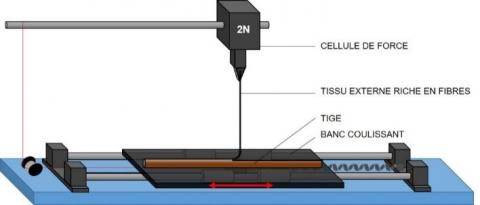
Les essais de pelage ont été effectués à température (23 ± 1 °C) et humidité relative (50 ± 1 %) contrôlées. Une machine d'essai de traction universelle MTS équipée d'une cellule de force d'une capacité de 2 N a été utilisée pour mesurer la force requise pour peler le tissu périphérique riche en fibres («tissus externes») du bois («tissus internes») des tiges. L'influence de la vitesse de déplacement a d'abord été étudiée. Avant pelage, des échantillons de 220 mm de long ont été prélevés à mi-hauteur de la plante (mesurant initialement entre 2 et 3 mètres de hauteur) et deux coupes longitudinales parallèles espacées de 1,5 mm ont été réalisées à l'aide de lames de rasoir. Une attention particulière a été apportée pour effectuer l’essai de pelage entre deux nœuds de la tige pour éviter toute perturbation. Les échantillons ont ensuite été fixés sur un support libre de déplacement horizontal à la même vitesse que le pelage du tissu extérieur de la tige à lieu verticalement, en maintenant un angle de 90° entre le tissu pelé et la tige pendant l’essai. La configuration expérimentale développée est illustrée en Figure 2. L'énergie de rupture moyenne, Gf, est calculée à partir de la courbe force/déplacement après stabilisation de la force, entre 5 et 15 mm.
Figure 2. Configuration expérimentale du test de pelage
2.4 Traction des tissus externes de la tige
Les tissus pelés ont été extraits après essais de pelage des tiges présentant un degré de rouissage différent. Les échantillons obtenus, d’une longueur de 70 mm et une largeur de 1,5 mm, ont été mesurés à l'aide d'un micromètre. La longueur de l'échantillon a été choisie pour garantir la caractérisation de la cohésion des faisceaux de fibres plutôt que les propriétés mécaniques des fibres elles-mêmes. Chaque échantillon a ensuite été fixé entre les mors de la machine de traction avec une longueur nominale de 50 mm. Les échantillons ont ensuite été testés à une vitesse de 50 mm.min-1, similaire à celle utilisée lors de l’essai de pelage afin de pouvoir comparer les résultats des deux essais. La courbe force/déplacement permet ensuite de calculer la rigidité (entre 0,5 % et 1,0 % de déformation), la contrainte et la déformation maximale du tissu externe contenant les fibres. L'énergie élastique dissipée lors du pelage pendant l’essais est ensuite calculée en supposant que la transition entre la bande non pelée et pelée, de déformation uniaxiale, se produit sur une longueur constante et que l'énergie de déformation élastique ne varie pas pendant l’essai de pelage.
2.5 Traction de fibres élémentaires
Des essais de traction sur fibres élémentaires ont été réalisés selon la norme NF T 25-501-2 qui prend en compte la complaisance du capteur de force. 60 fibres au minimum ont été testées, extraites de faisceaux issus des tiges ayant différents degrés de rouissage (R0, R1 et R2). Avant l’essai, les fibres unitaires ont été collées sur un cadre en papier pour obtenir une longueur de jauge fixe de 10 mm et conditionnées pendant 24 heures à une température et une humidité contrôlées à respectivement 23 °C et 50 %. Le diamètre de chaque fibre a été déterminé à l'aide d'un microscope optique, la valeur correspondant à la moyenne de six mesures effectuées le long de la fibre. Les échantillons ont ensuite été testés sur une machine d'essai de traction universelle MTS équipée d'un capteur de force de capacité 2 N avec une vitesse de 1 mm.min-1.
2.6 Microscopie électronique à balayage
Les échantillons ont été métallisés et recouverts d’une couche mince d'or de 15 nm et observés sous un microscope à balayage électronique Jeol JSM 6460LV.
2.7 Analyse statistique
Une analyse de variance à un facteur (one-way ANOVA + test post hoc de Tukey) a été effectuée. Les valeurs moyennes sont considérées comme statistiquement différentes si P <0,05.
3.1 État initial des échantillons
La Figure 3 montre une tige de chanvre non rouie (R0) après un test de pelage, et permet de visualiser le choix de la largeur de la bande pelée choisie (1,5 mm) pour assurer le pelage de la tige sur sa surface la plus plate. Cette géométrie permet de limiter les perturbations causées par la courbure transverse de la bande pelée qui pourraient être induites par la section irrégulière de la tige.
Les images de microscopie (cf. Figure 4.a) indiquent que le mécanisme de pelage se produit par propagation de fissure dans la région entre le tissu pelé et le reste de la tige, région où l’énergie nécessaire à la rupture des cellules est minimale. Cette région (illustrée sur le schéma d’une section de tige, Figure 4.b) est adjacente aux faisceaux de fibres et au bois. À l'échelle de la tige, elle se caractérise par une région tridimensionnelle contenant plus d'une couche moléculaire et, par conséquent, définie comme une interphase fibre/bois.
Figure 3. Image au microscope électronique à balayage d'une tige de chanvre non rouie après essai de pelage sachant que la largeur de la bande pelée est fixée pour éviter toutes contraintes de flexion transverse
La Figure 4.a de la tige pelée montre que le pelage se produit principalement par séparation du tissu externe (épiderme, parenchyme, fibres primaires, phloème secondaire) du bois de la tige (xylème secondaire). Pour une meilleure compréhension, la Figure 4.b montre un schéma d’une coupe transversale de tige de chanvre à mi-hauteur. Les fibres sont situées en périphérie de la tige et sont disposées en faisceaux [29]. La propagation des fissures se situe dans la région de la plante ayant la plus faible cohésion transverse, c'est-à-dire dans la région du phloème. À l'échelle microscopique, l’arrangement spécifique des cellules de cette région du phloème entraîne la propagation de fissures soit par séparation des parois cellulaires au niveau des lamelles mitoyennes, soit par rupture des parois cellulaires, ceci en fonction de la composition biochimique et de la densité des tissus [30-31]. Dans les plantes, les composés phénoliques, dont la lignine en est un groupe important, jouent un rôle prédominant dans la cohésion et la résistance mécanique des parois cellulaires [32]. La distribution spatiale de la lignine dans les tissus de la tige est complexe et inhomogène. Des études histologiques de coupe de tige révèlent une différence significative de teneur en lignine entre les tissus internes (phloème, cambium et xylème) et les tissus externes (épiderme, parenchyme et faisceaux de fibres) : dix fois plus élevée dans les tissus internes que dans les tissus externes [33]. Crônier et al. (2005) [34] a également montré l'absence de lignine par détection cytochimique dans les régions du phloème, qui semble être corrélée à la facilité de séparation entre les tissus riches en fibres du reste de la tige [34]. La région du phloème, au sein de laquelle la cohésion cellulaire est donc faible, peut être considérée comme une région prédominante pour la propagation de fissure lors du pelage. On peut donc supposer que cette région représente le maillon le plus faible pour ce type spécifique de chargement.
Figure 4. (a) Vue de côté d’une tige de chanvre non rouie pendant l’essai de pelage du tissu externe riche en fibres et b) Coupe schématique d'une tige de chanvre montrant l'organisation micro-structurelle sachant que la région en rouge représente l’interphase fibre/bois, au sein de laquelle le pelage à lieu
3.2 Comportement de la tige lors du pelage
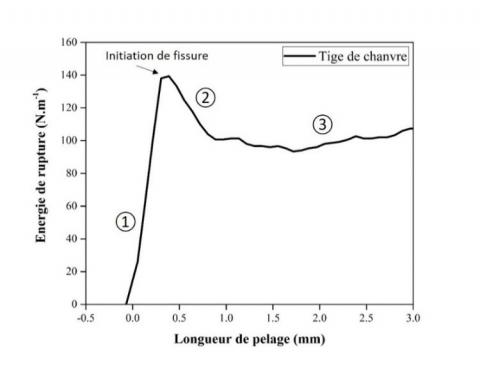
Figure 5. Comportement des tiges de chanvre lors de l'essai de pelage
L'énergie de rupture de l'interphase fibre/bois dans les tiges de chanvre a été évaluée par essais de pelage. La Figure 5 présente le comportement de pelage des tiges, montrant l'évolution de l'énergie de rupture en fonction de la distance de pelage. Au stade initial du pelage, une première augmentation de la force est linéaire de par la flexion élastique du tissu pelé avant initiation de la fissure (zone 1). Dans cette région de la courbe, les fissures sont stables jusqu'à ce qu'une force critique soit atteinte puis se propagent de façon stable au niveau de l'interphase entre le tissu externe riche en fibres et le xylème. L'énergie de rupture correspond à l'énergie nécessaire pour rompre la liaison interfaciale au niveau du front de pelage. Une fois la fissure initiée, une chute de l'énergie de rupture se produit en raison d’un déchargement élastique (zone 2) qui varie ensuite autour d'une valeur moyenne pendant le pelage continu du tissu pelé le long de la tige (zone 3).
3.3 Influence de la vitesse de pelage
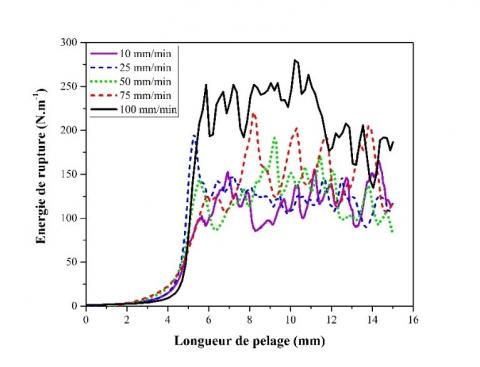
La force de pelage a été évaluée pour des échantillons de chanvre non rouis à 23 °C et avec un angle de 90° entre l'échantillon et la direction de la force appliquée (cf. Figure 2), sur une plage de vitesse de déplacement variant de 10 à 100 mm.min-1. Les résultats sont reportés sur la Figure 6 qui montre l'énergie de rupture, Gf, en fonction de la longueur de pelage le long de la tige. L'énergie de rupture est dépendant de la vitesse d'essai. Ce comportement n’est pas surprenant étant donné que la valeur de l’énergie de rupture reflète l’énergie plastique et viscoélastique dissipée localement dans la zone en amont du front de fissure, qui dépend également de la vitesse d’essai et de la température [35]. À faible vitesse (<100 mm.min-1), Gf tend vers une valeur constante autour de 140 ± 18 N.m-1, considérée comme la force de pelage à l'équilibre. Cette observation est en accord avec des études antérieures sur des stratifiés flexible [35], où l’énergie de rupture décroît avec la vitesse de pelage et approche une valeur mesurant directement la force de liaison moléculaire agissant sur l’interphase. Une vitesse de pelage de 50 mm.min-1 a donc été fixée pour les tests de pelage ultérieurs.
Figure 6. Évolution de l'énergie de rupture à l’interphase fibres/bois de tiges de chanvre non-rouies en fonction de la vitesse de pelage
3.4 Influence du rouissage : Approche multi-échelles
Interphase fibre/bois :
Des tiges de chanvre ayant un temps de rouissage variant de non-rouies (R0) à 17 jours (R1) et 37 jours (R2) ont été testées par essai de pelage. Le Tableau 1 indique la variation de l'énergie de rupture à l’interphase en fonction du degré de rouissage des tiges. L’énergie de rupture moyenne chute de 238 ± 99 Nm-1 à 117 ± 33 Nm-1, puis à 66 ± 39 Nm-1 pour R0, R1 et R2, respectivement. Ces résultats traduisent une réduction drastique de l’énergie absorbée lors du pelage du tissu externe pour des échantillons ayant un temps de rouissage plus longs. Cette diminution d’énergie est probablement due à une dégradation de l'interphase entre le tissu externe et le bois résultant du rouissage. En effet, l'activité enzymatique (principalement les polygalacturonases et les xylanases) présente dans les tiges au cours du rouissage est responsable de la dégradation des composants cellulaires et dépend de la composition chimique et de la pénétration des micro-organismes. Pendant le rouissage, les bactéries et/ou champignons se développent premièrement sur la surface externe et les extrémités ouvertes de la tige avant pénétration dans la région interne. La forte activité enzymatique entraîne une dégradation des lignines et des pectines entre les faisceaux de fibres, pour atteindre finalement l'interphase fibre/bois et dégrader les composants de cette région [36-38]. La décohésion des parois cellulaires se produit principalement dans les régions du phloème et du cambium, où les parois cellulaires sont plus minces et ont une plus faible teneur en lignine, connue pour inhiber la colonisation microbienne [15].
Tableau 1. Évolution de l’énergie de rupture moyenne en fonction du temps de rouissage des tiges de chanvre
|
Échantillons |
Temps de rouissage |
Énergie de rupture (N.m-1) |
|
R0 |
0 jour |
238 ± 99 |
|
R1 |
17 jours |
117 ± 33 |
|
R2 |
37 jours |
66 ± 39 |
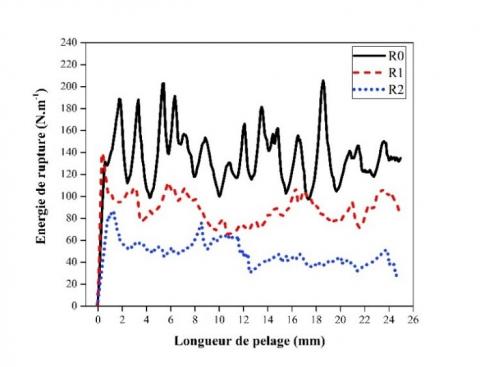
Figure 7. Évolution de l'énergie de rupture en fonction du degré de rouissage du chanvre
La Figure 7 montre l'évolution de l'énergie de rupture lors de la propagation de la fissure le long de la tige pour 3 échantillons présentant des diamètres équivalents de 5,1 ± 0,2 mm : R0, R1 et R2. En plus d’une diminution de l'énergie de rupture avec l’avancée du rouissage, une variation de cette énergie est présente pendant la phase de pelage après initiation de la fissure. Ce comportement démontre que le mécanisme de pelage implique la propagation de fissures instables et discontinues au niveau de l'interphase fibre/bois. Ce phénomène est plus ou moins marqué entre les échantillons, avec une atténuation des fluctuations vers des degrés de rouissage plus élevés.
Les mécanismes de rupture mis en jeu sont analysés en observant la morphologie de la surface de pelage des nouvelles surfaces créées lors de l’essai. Des images de microscope électronique à balayage (cf. Figure 8) montrent les surfaces du tissu pelé et du bois, initialement en contact avant essai, pour R0, R1 et R2. À l'échelle microscopique, le pelage des tissus riches en fibres se produit par désintégration, ou rupture cohésive, des cellules au niveau de l'interphase fibre/bois pour l'échantillon non roui, laissant une surface de pelage rugueuse. Ainsi, de grandes fluctuations de l'énergie de rupture sont observées lors des essais, chaque cellule agissant comme un défaut perturbant la propagation de la fissure. Plus le degré de rouissage est élevé, plus l'interphase fibre/bois est dégradée et donc une évolution significative du rapport pectine/lignine peut être supposée. Les lignines, composants plus rigides et plus plastifiés que les pectines, deviennent prédominants conduisant à une rupture adhésive des cellules, provoquée par la séparation ou pelage des faisceaux de fibres au niveau de l'interphase [4, 21]. Il peut également être supposé que la dégradation des composants le long des fibres provoquée par le rouissage créée une région «affaiblie» en périphérie des faisceaux de fibres. Dans le cas des échantillons R1, rouissage intermédiaire, les deux types de surface sont observés, entraînant un mécanisme de pelage mixte, impliquant pelage et rupture des cellules, ce qui conduit à une rupture partiellement cohésive. Cette variation de mécanisme de pelage a également été décrite par Ashby (1985) [30], qui montre que la ténacité du bois dépend de sa structure, ce qui conduit à des valeurs plus élevées lorsque des phénomènes de rupture des parois cellulaires se produisent, comparable aux échantillons non-rouis dans cette étude.
Figure 8. Images au microscope électronique à balayage montrant les surfaces de pelage initialement en contact avant essai avec les schémas représentant le comportement à la rupture pour les échantillons R0, R1 et R2
Tissu externe riche en fibres :
Ajouté à la dégradation de l’interphase fibres/bois, l’étape de rouissage affecte également la cohésion des faisceaux de fibres. Des essais mécaniques ont été effectués sur des tissus externes (composés d’épiderme, de faisceaux de fibres et de phloème) extraits de tiges non rouis et de tiges rouies à 37 jours. Le Tableau 2 montre l'impact du degré de rouissage sur les propriétés de traction de ces tissus, ainsi que les résultats des tests ANOVA comparant les différences statistiques du module tangent, la contrainte et la déformation maximale entre chaque degré de rouissage. Très peu de variations sont observées en terme de propriétés mécaniques des faisceaux de fibres entre les échantillons non rouis et rouis. Pour les échantillons R2, le module tangent est environ 7 % plus élevé que pour les échantillons R0, 3 % et 9 % plus faible pour la contrainte et la déformation maximale. Néanmoins, les valeurs ne sont pas statistiquement différentes selon les tests ANOVA.
Tableau 2. Propriétés mécaniques en traction du tissu externe riche en fibres pour les échantillons non rouis (R0) et rouis (R2), ainsi que les résultats des tests statistiques ANOVA. E.T = écart-type ; E = module tangent moyen ; σ = contrainte maximale ; ε = déformation maximale
|
|
Non-roui R0 |
Roui R2 |
Statistique ANOVA |
|
|
|
Moyenne ± E.T |
Moyenne ± E.T |
Prob>F |
α |
|
E (GPa) |
9,56± 4.29 |
10,27± 1,83 |
0,7416 |
0,05 |
|
σ (MPa) |
177,0± 79,6 |
172,4± 76,7 |
0,9290 |
0,05 |
|
ε (%) |
2,03± 0,04 |
1,85± 0,60 |
0,5061 |
0,05 |
Les propriétés mécaniques des tissus externes peuvent difficilement être comparées aux données de la littérature en raison de la nature de l'échantillon (présence d'épiderme, de parenchyme cortical et de faisceaux de fibres). Malgré des propriétés mécaniques de traction similaires entre R0 et R2, le comportement à rupture de ces tissus est différent. La Figure 9 montre des images de microscopie électronique à balayage des faciès de rupture des tissus externes ainsi que des schémas illustrant le comportement à rupture après essai de traction. Les échantillons non rouis, dans lesquels les fibres sont liées entre elles par une interphase pectique, présentent une rupture fragile des fibres individuelles, accompagnée de mécanismes de glissement dus à une forte cohésion entre les fibres. Les propriétés de résistance à la traction dépendent donc à la fois des propriétés mécaniques des fibres individuelles composant les faisceaux et d’un effort de cisaillement au sein des lamelles mitoyennes. Dans le cas des échantillons rouis, une décohésion peut être observée dans les faisceaux de fibres en raison de la dégradation des lamelles mitoyennes résultant du rouissage. Les propriétés mécaniques des échantillons rouis dépendent dans ce cas principalement des propriétés de cohésion interfaciale fibres/fibres au sein des faisceaux. Cependant, la structure complexe de ces tissus externes, en particulier l’influence des lamelles mitoyennes, ne permet pas d’observer des différences évidentes en terme de propriétés mécaniques des faisceaux de fibres avant et après rouissage.
Figure 9. Images de microscopie électronique à balayage et schémas représentant les faciès de rupture des tissus externes des tiges de chanvre non rouies (R0) en haut et rouies (R2) en bas
Issue de ces mesures de propriétés mécaniques, et plus précisément de la déformation longitudinale des tissus externes, l’énergie de rupture obtenue ne représente pas une énergie «vraie» à l’interphase définie comme la force nécessaire pour peler les tissus riches en fibres du reste de la tige. En effet, la résistance à la rupture de l’interphase n'est pas la seule source de dissipation d'énergie dans ce système. Pour déterminer la valeur d’énergie de rupture réelle, il est nécessaire d’appliquer des corrections prenant en compte l’existence d’une énergie dissipée pendant la déformation longitudinale du tissu externe [25]. Compte tenu de la faible épaisseur de celui-ci, l'absence de flexion plastique au niveau du front de fissure peut être supposée [35]. L'énergie de rupture s’exprime donc par deux termes, l'énergie de rupture au front de fissure et l'énergie dissipée pour extension du tissu externe (équation 2) :
$G_{f}=\frac{F}{b}(1-\cos \theta)+\frac{1}{2 . E . h}\left(\frac{F}{b}\right)^{2}$ (2)
où Gf est l'énergie de rupture totale, F la force de pelage, b la largeur de tissu pelé (1.5 mm), θ l'angle de pelage (90°), E le module tangent du tissu externe et h son épaisseur. Les valeurs mesurées de l'énergie de rupture, l'énergie dissipée due à l'expansion et au pelage du tissu externe, ainsi que l'énergie de pelage calculée sont donnés dans le Tableau 3.
Tableau 3. Énergie de rupture mesurée Gf des tiges de chanvre non rouies (R0) et rouis (R2) au cours de l’essai de pelage avec l'énergie dissipée lors de l'extension du tissu externe ainsi que l'énergie de pelage «réelle»
|
|
|
Énergie de rupture dissipée |
|
|
|
Énergie de Rupture totale Gf (N.m-1) |
Pour l’extension du tissu externe (N.m-1) |
Pour le pelage du tissu externe (N.m-1) |
|
|
Moyenne ± E.T |
Moyenne ± E.T |
Moyenne ± E.T |
|
R0 |
233,3 ± 111,2 |
34,9 ± 17,6 |
198,4 ± 73,7 |
|
R2 |
50,0 ± 17,0 |
0,9 ± 0,5 |
49,1 ± 16,5 |
Les valeurs calculées à partir de l'énergie correspondant au pelage du tissu externe sont nettement inférieures à l'énergie de rupture mesurée Gf. L'énergie de pelage corrigée atteint 198,4 N.m-1 pour les tiges non rouies et 49,1 N.m-1 pour les tiges rouies. Également, l'énergie dissipée par extension du tissu externe dépend du degré de rouissage et représente 14.9 % de l'énergie de rupture totale pour R0 et 1.8 % pour R2. Ce comportement peut être lié à la déformation à rupture plus élevée observée lors des essais de traction des tissus externes non-rouis. En d'autres termes, le rouissage induit une dégradation des lamelles mitoyennes et la cohésion entre les fibres, réduisant le transfert de charge dans les zones inter-fibres.
Fibres élémentaires :
Cette dernière section se concentre sur l’impact du rouissage sur les propriétés mécaniques des fibres élémentaires. Les valeurs du module apparent, de la contrainte et déformation maximales sont indiquées dans le Tableau 4, ainsi que les diamètres moyens des fibres individuelles testées.
Tableau 4. Propriétés mécaniques des fibres élémentaires présentant un rouissage différent. E.T = écart type ; E = module apparent moyen ; σ = contrainte maximale ; ε = déformation maximale
|
|
Non-rouies R0 |
Rouies R2 |
Statistique ANOVA |
|
|
|
Moyenne± E.T |
Moyenne± E.T |
Prob > F |
α |
|
E(GPa) |
18,26± 13,07 |
24,22± 16,64 |
0,0328 |
0,05 |
|
σ(MPa) |
406,7± 255,7 |
432,8± 375,6 |
0,6666 |
0,05 |
|
ε(%) |
3,52± 1,98 |
2,87± 1,65 |
0,4589 |
0,05 |
Le diamètre des fibres décroît progressivement avec l’avancée du rouissage (33,5 ± 10,5 µm pour les fibres non rouies et 29,4 ± 7,7 µm pour les fibres rouies). Les tests ANOVA permettent de confirmer que les diamètres des fibres R0 et R2 sont statistiquement différents (Prob> F = 0,0146 avec α = 0.05). Cette légère diminution du diamètre moyen est associée à l'élimination des composants de surface des fibres tels que la pectine et la lignine présentes dans les lamelles mitoyennes sous l'action du rouissage.
Le module apparent moyen des échantillons non rouis atteint une valeur moyenne de 18,26 GPa, une contrainte maximale de 406,7 MPa et une déformation maximale de 3,52 %. Ces valeurs sont proches des données de la littérature de plusieurs travaux [4, 39-41]. Le rouissage induit une variation des propriétés de traction des fibres élémentaires. Un degré de rouissage plus élevé (R2) entraîne une augmentation de 6 % et 33 % de la contrainte maximale et du module apparent, respectivement, et une diminution de 18 % de la déformation maximale. Une analyse statistique confirme une différence de module tangent entre les échantillons non rouis et rouis, contrairement aux valeurs de contrainte et de déformation qui sont considérées comme statistiquement non différents. La grande dispersion des propriétés en traction s'explique principalement par l'incertitude de mesure sur la section transversale des fibres et par l'hétérogénéité du rouissage. Une étude antérieure montrant l'effet du rouissage sur les propriétés de traction des fibres de lin a révélé une tendance similaire avec une augmentation du module tangent et de la contrainte maximale [42]. L’élévation des performances en traction des fibres rouies proviennent de multiples phénomènes complexes mais peuvent cependant s'expliquer par la dégradation de matériau peu performant mécaniquement que sont les lamelles mitoyennes et par la facilité de défibrage des fibres engendrant potentiellement moins de defaults internes.
De nos jours, les producteurs de chanvre évaluent la facilité d'extraction des faisceaux ainsi que l'état de biodégradation par manipulation de la tige. L'essai de pelage développé dans cette étude aborde cette évaluation de manière standardisée en donnant des informations sur plusieurs aspects : le comportement mécanique et l'énergie nécessaire lors du pelage des tiges mettant en évidence une région prédominante pour la propagation de fissure. De plus, des essais mécaniques sur tissus externes (principalement constitués de faisceaux de fibres) ont été effectués et sont un moyen de vérifier la non-dégradation des fibres due à un éventuel sur-rouissage. Des vérifications similaires sont effectuées dans le cas du lin textile en utilisant des méthodes organoleptiques qui ne donnent aucune information sur la facilité d'individualiser les faisceaux de fibres.
Ce travail montre tout d'abord l'impact du degré de rouissage de la tige sur le comportement à l'interphase fibre/bois, avec une réduction de 75 % de l'énergie de rupture lors du pelage du tissu fibreux, permettant ainsi une compréhension plus avancée des mécanismes de rouissage à l'échelle des tissus de la tige. Grâce à une analyse de l'évolution de l'énergie de pelage et une étude morphologique de la zone de pelage par MEB, nous mettons en évidence l'impact du rouissage sur la cohésion et la microstructure de la tige ; une dégradation des composants riches en pectine est supposée, induisant de fortes modifications dans les mécanismes de rupture au sein des tissus de la tige. Combinée à des essais de traction sur faisceaux de fibres, cette étude montre la capacité du rouissage à réduire l'énergie de rupture au niveau de l'interphase fibre/bois et à réduire la cohésion des fibres au sein des faisceaux. Il a donc été démontré que des essais de pelage peuvent être utilisés pour suivre l’avancée du rouissage facilitant l’extraction des faisceaux de fibres des tiges. À l’échelle de la fibre, cela pourrait permettre de limiter la création de défauts et d’obtenir une matière première qualité supérieure. Les essais de traction des fibres élémentaires confirment cette hypothèse en montrant des performances mécaniques améliorées (augmentation de 33 % du module de traction) entre les échantillons non rouis et rouis.
Cette étude offre une méthode fiable pour déterminer le degré de rouissage des plantes, afin de définir le degré de rouissage optimal pour faciliter l'extraction des fibres, conduisant à des fibres de meilleure qualité et, par conséquent, à la production de matériaux composites plus performants. Cependant, des recherches plus approfondies sont nécessaires pour comprendre l’impact du rouissage à l’échelle des fibres élémentaires (propriétés mécaniques, morphologie, rugosité de surface) pour analyser plus précisément les performances des faisceaux de fibres ainsi que l’impact du rouissage sur la composition chimique et de la cristallinité de la cellulose.
Les auteurs remercient l'ADEME pour son soutien financier au projet et les membres du projet RIGHTLAB, à savoir FRD, La Chanvrière et l'unité FARE de l'INRA.
[1] Beckermann, G.W., Pickering, K.L. (2008). Engineering and evaluation of hemp fibre reinforced polypropylene composites: Fibre treatment and matrix modification. Composites Part A: Applied Science and Manufacturing, 39(6): 979-988. https://doi.org/10.1016/j.compositesa.2008.03.010
[2] Pickering, K.L., Beckermann, G.W., Alam, S.N., Foreman, N.J. (2007). Optimising industrial hemp fibre for composites. Composites Part A: Applied Science and Manufacturing, 38(2): 461-468. https://doi.org/10.1016/j.compositesa.2006.02.020
[3] Placet, V. (2009). Characterization of the thermo-mechanical behaviour of Hemp fibres intended for the manufacturing of high performance composites. Composites Part A: Applied Science and Manufacturing, 40(8): 1111-1118. https://doi.org/10.1016/j.compositesa.2009.04.031
[4] Placet, V., Day, A., Beaugrand, J. (2017). The influence of unintended field retting on the physicochemical and mechanical properties of industrial hemp bast fibres. Journal of Materials Science, 52(10): 5759-5777. https://doi.org/10.1007/s10853-017-0811-5
[5] Lühr, C., Pecenka, R., Budde, J., Hoffmann, T., Gusovius, H.J. (2018). Comparative investigations of fibreboards resulting from selected hemp varieties. Industrial Crops and Products, 118: 81-94. https://doi.org/10.1016/j.indcrop.2018.03.031
[6] Müssig, J., Amaducci, S. (2018). Scanner based image analysis to characterise the influence of agronomic factors on hemp (Cannabis sativa L.) fibre width. Industrial Crops and Products, 113: 28-37. https://doi.org/10.1016/j.indcrop.2017.12.059
[7] Wang, S., Gusovius, H.J., Lühr, C., Musio, S., Uhrlaub, B., Amaducci, S., Müssig, J. (2018). Assessment system to characterise and compare different hemp varieties based on a developed lab-scaled decortication system. Industrial Crops and Products, 117: 159-168. https://doi.org/10.1016/j.indcrop.2018.02.083
[8] Liu, M., Fernando, D., Daniel, G., Madsen, B., Meyer, A.S., Ale, M.T., Thygesen, A. (2015a). Effect of harvest time and field retting duration on the chemical composition, morphology and mechanical properties of hemp fibers. Industrial Crops and Products, 69: 29-39. https://doi.org/10.1016/j.indcrop.2015.02.010
[9] Müssig, J., Martens, R. (2003). Quality aspects in hemp fibre production — influence of cultivation, harvesting and retting. Journal of Industrial Hemp, 8(1): 11-32. https://doi.org/10.1300/J237v08n01_03
[10] Tahir, P., Ahmed, A., SaifulAzry, S., Ahmed, Z. (2011). Retting process of some bast plant fibres and its effect on fibre quality: A review. BioResources, 6(4): 5260-5281. https://doi.org/10.15376/biores.6.4.5260-5281
[11] Aslan, M., Mehmood, S., Madsen, B., Goutianos, S. (2010). The effect of processing on defects and tensile strength of single flax fibres. In: 14th European Conference on Composite Materials.
[12] Bos, H.L., Van Den Oever, M.J.A., Peters, O.C.J.J. (2002). Tensile and compressive properties of flax fibres for natural fibre reinforced composites. Journal of Materials Science, 37(8): 1683-1692. https://doi.org/10.1023/A:1014925621252
[13] Hernandez-Estrada, A., Gusovius, H.J., Müssig, J., Hughes, M. (2016). Assessing the susceptibility of hemp fibre to the formation of dislocations during processing. Industrial Crops and Products, 85: 382-388. https://doi.org/10.1016/j.indcrop.2016.01.006
[14] Thygesen, A., Madsen, B., Bjerre, A.B., Lilholt, H. (2011). Cellulosic fibers: Effect of processing on fiber bundle strength. Journal of Natural Fibers, 8(3): 161-175. https://doi.org/10.1080/15440478.2011.602236
[15] Akin, D.E., Gamble, G.R., Morrison III, W.H., Rigsby, L.L., Dodd, R.B. (1996). Chemical and structural analysis of fibre and core tissues from flax. Journal of the Science of Food and Agriculture, 72(2): 155-165. https://doi.org/10.1002/(SICI)1097-0010(199610)72:2<155::AID-JSFA636>3.0.CO;2-X
[16] Meijer, W.J.M., Vertregt, N., Rutgers, B., van de Waart, M. (1995). The pectin content as a measure of the retting and rettability of flax. Industrial Crops and Products, 4(4): 273-284. https://doi.org/10.1016/0926-6690(95)00041-0
[17] Saleem, Z., Rennebaum, H., Pudel, F., Grimm, E (2008). Treating bast fibres with pectinase improves mechanical characteristics of reinforced thermoplastic composites. Composites Science and Technology, 68(2): 471-476. https://doi.org/10.1016/j.compscitech.2007.06.005
[18] Djemiel, C., Grec, S., Hawkins, S. (2017). Characterization of bacterial and fungal community dynamics by high-throughput sequencing (HTS) metabarcoding during flax dew-retting. Front. Microbiol., 8: 1-16. https://doi.org/10.3389/fmicb.2017.02052
[19] Li, Y., Pickering, K.L., Farrell, R.L. (2009). Analysis of green hemp fibre reinforced composites using bag retting and white rot fungal treatments. Industrial Crops and Products, 29(2-3): 420-426. https://doi.org/10.1016/j.indcrop.2008.08.005
[20] Liu, M., Thygesen, A., Summerscales, J., Meyer, A.S. (2017b). Targeted pre-treatment of hemp bast fibres for optimal performance in biocomposite materials: A review. Industrial Crops and Products, 108: 660-683. https://doi.org/10.1016/j.indcrop.2017.07.027
[21] Mazian, B., Bergeret, A., Benezet, J.C., Malhautier, L. (2018). Influence of field retting duration on the biochemical, microstructural, thermal and mechanical properties of hemp fibres harvested at the beginning of flowering. Industrial Crops and Products, 116: 170-181. https://doi.org/10.1016/j.indcrop.2018.02.062
[22] Gardon, J.L. (1963). Peel adhesion. I. Some phenomenological aspects of the test. J. Appl. Polym. Sci., 7(2): 625-641. https://doi.org/10.1002/app.1963.070070219
[23] Gent, A.N., Petrich, R.P. (1969). Adhesion of viscoelastic materials to rigid substrates. Proc. R. Soc. A Math. Phys. Eng. Sci., 310(1502): 433-448. https://doi.org/10.1098/rspa.1969.0085
[24] Kaelble, D. (1964). Theory and analysis of peel adhesion: Rate-temperature dependence of viscoelastic interlayers. Journal of Colloid Science, 19(5): 413-424. https://doi.org/10.1016/0095-8522(64)90041-8
[25] Kendall, K. (1973). The shapes of peeling solid films. The Journal of Adhesion, 5(2): 105-117. https://doi.org/10.1080/00218467308075015
[26] Toury-Triboulot, M.., Triboulot, P. (2012). Matériaux bois - Structure et caractéristiques. Tech. l’ingénieur c925.
[27] Wang, L., Lu, Z., Zhao, G. (2003). Wood fracture pattern during the water adsorption process. Holzforschung, 57(6): 639-643. https://doi.org/10.1515/HF.2003.096
[28] Booth, I., Goodman, A.M., Grishanov, S.A., Harwood, R.J. (2004). A mechanical investigation of the retting process in dew-retted hemp (Cannabis sativa). Annals of Applied Biology, 145(1): 51-58. https://doi.org/10.1111/j.1744-7348.2004.tb00358.x
[29] Réquilé, S., Goudenhooft, C., Bourmaud, A., Le Duigou, A., Baley, C. (2018). Exploring the link between flexural behaviour of hemp and flax stems and fibre stiffness. Industrial Crops and Products, 113: 179-186. https://doi.org/10.1016/j.indcrop.2018.01.035
[30] Ashby, M.F., Easterling, K.E., Harrysson, R., Maiti, S.K. (1985). The fracture and toughness of woods. Proceedings of the Royal Society a Mathematical, Physical and Engineering Sciences, 398(1815): 261-280. https://doi.org/10.1098/rspa.1985.0034
[31] Bardet, S., Beauchene, J., Thibaut, B. (2003). Influence of basic density and temperature on mechanical properties perpendicular to grain of ten wood tropical species. Annals of Forest Science, 60(1): 49-59. https://doi.org/10.1051/forest:2002073
[32] Girault, R., His, I., Andeme-onzighi, C., Driouich, A., Morvan, C. (2000). Identification and partial characterization of proteins and proteoglycans encrusting the secondary cell walls of flax fibres. Planta, 211(2): 256-264. https://doi.org/10.1007/s004250000
[33] Liu, M., Fernando, D., Meyer, A.S., Madsen, B., Daniel, G., Thygesen, A. (2015b). Characterization and biological depectinization of hemp fibers originating from different stem sections. Industrial Crops and Products, 76: 880-891. https://doi.org/10.1016/j.indcrop.2015.07.046
[34] Crônier, D., Monties, B., Chabbert, B. (2005). Structure and chemical composition of bast fibers isolated from developing hemp stem. J. Agric. Food Chem., 53: 8279-8289. https://doi.org/10.1021/jf051253k
[35] Kinloch, A.J., Lau, C.C., Williams, J.G. (1994). The peeling of flexible laminates. International Journal of Fracture, 66(1): 45-70. https://doi.org/10.1007/BF00012635
[36] Brown, A.E., Sharma, H.S.S., Black, D.L.R. (1986). Relationship between pectin content of stems of flax cultivars, fungal cell wall-degrading enzymes and pre-harvest retting. Annals of Applied Biology, 109(2): 345-351. https://doi.org/10.1111/j.1744-7348.1986.tb05326.x
[37] Liu, M., Ale, M.T., Kołaczkowski, B., Fernando, D., Daniel, G., Meyer, A.S., Thygesen, A. (2017a). Comparison of traditional field retting and Phlebia radiata Cel 26 retting of hemp fibres for fibre-reinforced composites. AMB Express. https://doi.org/10.1186/s13568-017-0355-8
[38] Ribeiro, A., Pochart, P., Day, A., Mennuni, S., Bono, P., Baret, J.L., Spadoni, J.L., Mangin, I. (2015). Microbial diversity observed during hemp retting. Applied Microbiology and Biotechnology, 99(10): 4471-4484. https://doi.org/10.1007/s00253-014-6356-5
[39] Duval, A., Bourmaud, A., Augier, L., Baley, C. (2011). Influence of the sampling area of the stem on the mechanical properties of hemp fibers. Materials Letters, 65(4): 797-800. https://doi.org/10.1016/j.matlet.2010.11.053
[40] Liu, M., Meyer, A.S., Fernando, D., Silva, D.A.S., Daniel, G., Thygesen, A. (2016). Effect of pectin and hemicellulose removal from hemp fibres on the mechanical properties of unidirectional hemp/epoxy composites. Composites Part A: Applied Science and Manufacturing, 90: 724-735. https://doi.org/10.1016/j.compositesa.2016.08.037
[41] Placet, V., Trivaudey, F., Cisse, O., Gucheret-Retel, V., Boubakar, M.L. (2012). Diameter dependence of the apparent tensile modulus of hemp fibres: A morphological, structural or ultrastructural effect? Composites Part A: Applied Science and Manufacturing, 43(2): 275-287. https://doi.org/10.1016/j.compositesa.2011.10.019
[42] Martin, N., Mouret, N., Davies, P., Baley, C. (2013). Influence of the degree of retting of flax fibers on the tensile properties of single fibers and short fiber/polypropylene composites. Industrial Crops and Products, 49: 755-767. https://doi.org/10.1016/j.indcrop.2013.06.012