Swelling of Plant Fibers under Hygro/Hydrothermal Conditions: Determination of Hygro/Hydroexpansion Coefficients
Processus de Gonflement des Fibres Végétales en Conditions Hygro/Hydrothermiques : Détermination des Coefficients d’Hygro/Hydroexpansion

William Garat | Nicolas Le Moigne* | Stéphane Corn | Johnny Beaugrand | Anne Bergeret
OPEN ACCESS
The mechanical behavior of biocomposites and their durability are influenced by the intrinsic properties of the natural fiber reinforcements, especially their high sensitivity to hygro/hydrothermal conditions. To better understand these phenomena, the effect of humidity conditions on the moisture and dimensional variations of natural fiber bundles from several botanical origins with contrasting biochemical and structural characteristics was studied. Results evidenced differences in water uptake and swelling as a function of plant species and allowed to determine hygro/hydroexpansion coefficients of fiber bundles, thus opening interesting perspectives for the predictive modeling of “in-service” mechanical behavior of biocomposites.
RÉSUMÉ :
Le comportement mécanique des biocomposites et leur durabilité sont influencés par les propriétés intrinsèques des renforts végétaux, en particulier par leur forte sensibilité aux conditions hygro/hydrothermiques. Afin de mieux appréhender ces phénomènes, l’effet des conditions d’humidité sur la teneur en eau et les variations dimensionnelles de faisceaux de fibres végétales d’origines botaniques aux caractéristiques biochimiques et structurelles contrastées a été étudié. Les résultats ont mis en évidence des reprises en eau et des processus de gonflement différenciés en fonction des espèces végétales et ont permis la détermination des coefficients d’hygro/hydroexpansion des faisceaux de fibres, ouvrant ainsi des perspectives intéressantes pour la modélisation prédictive du comportement mécanique « en service » des biocomposites.
natural fibers, swelling, humidity, hygro/hydroexpansion coefficient
Mots-clés :
fibres végétales, gonflement, humidité, coefficient d’hygro/hydroexpansion
Au cours de leur utilisation, les matériaux composites peuvent être soumis à diverses sollicitations environnementales comme l’humidité, la température ou le rayonnement UV. Cette exposition aux conditions environnementales est la cause première de leur vieillissement prématuré. Ces processus de vieillissement demandent une attention toute particulière afin d’évaluer le comportement évolutif de ces matériaux composites en service, et particulièrement ceux des biocomposites renforcés par des fibres végétales. En effet, la stabilité dimensionnelle et le comportement mécanique des fibres végétales sont fortement influencés par les conditions hygro/hydrothermiques.
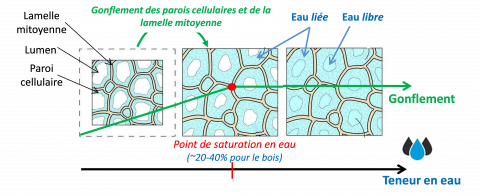
En fonction des conditions hygro/hydrothermiques environnantes, les fibres végétales peuvent contenir de l’eau sous deux formes : (i) l’eau liée dans les parois cellulaires et la lamelle mitoyenne, via la formation de liaisons hydrogènes avec les groupes hydroxyles -OH des différents biopolymères des parois [1, 2] et (ii) l’eau libre qui se trouve dans les micropores des parois cellulaires, remplissant les vides (lumens) et retenue par les forces capillaires. La Figure 1 représente schématiquement les processus de gonflement des cellules du bois en fonction de la teneur en eau. L’eau en phase vapeur pénètre et est absorbée dans les parois cellulaires et la lamelle mitoyenne via la formation de liaisons hydrogènes, provoquant ainsi un gonflement important des cellules. A une certaine teneur en eau, qui dépend de l’espèce végétale considérée, l’eau liée sature les parois cellulaires et la lamelle mitoyenne : le point de saturation en eau des cellules végétales est alors atteint, ~20-40 % pour le bois [3]. Enfin, les cavités cellulaires (lumens et porosités) sont remplies par l’eau libre jusqu’à saturation totale.
Plusieurs techniques de mesure permettent de quantifier la teneur en eau dans les fibres végétales en fonction de l’humidité relative (HR). On retiendra en particulier : (i) la DVS (Dynamic Vapor Sorption), qui est une technique gravimétrique mesurant la quantité d’eau absorbée par un échantillon en fonction de l’humidité relative d’équilibre (ou activité de l’eau aw = P/P0 avec P la pression de vapeur d’eau à la surface du substrat considéré et P0 la pression de vapeur saturante de l’eau pure). Plusieurs auteurs ont utilisé cette technique pour tracer les isothermes de sorption d’eau à partir d’un échantillon sec, ou les isothermes de désorption à partir d’un échantillon saturé en eau [4-6] ; (ii) la WRV (Water Retention Value) qui est une méthode de centrifugation et de pesée permettant de déterminer la valeur de rétention d’eau des fibres végétales après immersion. La WRV est déterminée selon la norme ISO 23714 [7].
Figure 1. Processus de gonflement des cellules du bois (adapté de [8])
Les processus de sorption d’eau et de gonflement des fibres végétales sont complexes du fait de leurs particularités biochimiques, structurelles et morphologiques. En ce sens, on notera que ces processus sont influencés par plusieurs facteurs, comme la dimension des lumens, l’angle microfibrillaire (MFA) [9], le taux de cristallinité de la cellulose, l’accessibilité aux fonctions hydroxyles, la teneur en polysaccharides/biopolymères amorphes et leur nature hydrophile/hydrophobe. Dans le cas des substrats (ligno-)cellulosiques, on constate la plupart du temps une forte hystérésis entre les isothermes de sorption et de désorption. Les origines de ce phénomène sont encore débattues dans la littérature et bien qu’il n’y ait pas encore de consensus, plusieurs auteurs l’attribuent à des processus de plastification et de relaxation des biopolymères amorphes présents dans les parois cellulaires [10, 11] en lien avec leur température de transition vitreuse. Par ailleurs, Karmaker et al. [12] ont mis en évidence l’hétérogénéité du gonflement de la section transversale des faisceaux de jute dans l’eau. Des faisceaux de jute secs ont été comparés aux mêmes faisceaux immergés deux semaines dans l’eau. Les observations en microscopie optique des variations dimensionnelles de plusieurs sections le long des faisceaux ont montré que le gonflement maximum se produisait dans les régions des faisceaux où les dimensions transverses étaient les plus faibles. Selon les auteurs, dans ces zones plus minces, les fibres élémentaires seraient plus compactées de sorte que lors de la sorption d’eau au sein du faisceau, le gonflement de chaque fibre élémentaire provoque une plus forte augmentation du diamètre.
L’ensemble de ces travaux met en évidence l’intérêt de mesurer de façon précise le gonflement des fibres végétales en lien avec leur teneur en eau. Ces données sont essentielles pour mieux décrire et comprendre les processus de gonflement des fibres végétales qui influencent directement la mise en œuvre/forme et le comportement en service des biocomposites. On notera en particulier que le gonflement des fibres peut induire un processus de frettage à l’interface fibre/matrice et contribuer à une amélioration de l’adhérence interfaciale [13], mais également générer des niveaux de contrainte susceptibles d'endommager localement la matrice et entraîner ainsi la dégradation du composite [14].
Le but de ce travail est de caractériser, dans des conditions hygro/hydrothermiques variables et contrôlées (20 %, 50 %, 73 % HR et immersion), la teneur en eau et les variations dimensionnelles de faisceaux de fibres végétales d’origines botaniques aux caractéristiques biochimiques et structurelles contrastées. Une technique de mesure de la section transversale par balayage laser automatisé [15] conduite soit dans une chambre climatique, soit dans une cellule d'immersion, a permis d'évaluer les variations des dimensions transversales des faisceaux de fibres en lien avec leurs teneurs en eau déterminée par des mesures de DVS et WRV. Ces analyses conduisent ainsi à déterminer les coefficients d’hygro/hydroexpansion des différents faisceaux de fibres végétales étudiées.
2.1 Sélection des faisceaux de fibres végétales
Des faisceaux de fibres végétales ont été extraits de cinq espèces phylogénétiquement différentes ayant des fonctions dissemblables dans la plante. Ces lots de faisceaux de fibres présentent des caractéristiques morphométriques [15], des microstructures et des compositions biochimiques contrastées. Trois d'entre eux appartiennent au clade des dicotylédones et sont communément appelés «fibres libériennes». Ces fibres sont faiblement lignifiées (< 5 %), et ont une fonction de support structurel de la tige dans la plante : le lin Linum usitatissimum (Alizée, Grandvilliers, Picardie, 2012), le chanvre Canabis sativa (Fédora 17, Champagne Ardenne, 2012) en culture annuelle de fibres (après rouissage) et l'ortie Urtica dioica (2014, Lorraine) comme plante herbacée pérenne. Les deux autres espèces font partie du clade des monocotylédones et sont vivaces. Ces fibres sont fortement lignifiées (> 9 %) : le sisal Agave sisalana (FRD, Fibre Recherche Développement®, Troyes) provenant de la feuille d'agave et le palmier Phoenix dactylifera (Al-Ahsa, Arabie Saoudite) présent sous forme de tapis de gaine foliaire entourant la tige. Les compositions biochimiques et MFA de ces différents lots sont données en Tableau 1.
Tableau 1. Teneur massique en polysaccharides cellulosiques et non-cellulosiques (par analyse HPLC), en lignine (méthode de Klason) et MFA (tirés de la littérature [16, 17, 18]) pour les cinq types de faisceaux de fibres végétales considérées
|
Espèce végétale |
Cellulose (%) |
Hémicellulose (%) |
Lignine (%) |
Autres (pectines...) (%) |
MFA (°) |
|
Palmier |
45,1 ± 3,4 |
27,7 ± 4,9 |
16,2 ± 0,3 |
10 |
40-47 |
|
Sisal |
60,1 ± 1,2 |
23,3 ± 2,1 |
9,3 ± 0,1 |
8 |
18-22 |
|
Chanvre |
68,8 ± 1,3 |
9,8 ± 1,8 |
4,7 ± 0,3 |
17 |
6-11 |
|
Lin |
64,9 ± 2,9 |
9,9 ± 3,9 |
3,7 ± 0,3 |
21 |
6-10 |
|
Ortie |
51,7 ± 0,9 |
9,9 ± 1,2 |
4,6 ±0,2 |
34 |
7,5 |
2.2 Caractérisation des faisceaux de fibres végétales en conditions hygro/hydrothermiques contrôlées
2.2.1 Mesure de la teneur en eau
La détermination des isothermes d’adsorption d’eau et des teneurs en eau à HR donnée pour les différentes fibres végétales a été réalisée par analyse DVS (Dynamic Vapor Sorption). Ces essais ont été réalisés sur une plage d’humidité relative comprise entre 0 et 90 % HR, par intervalles HR de 10 % avec une quantité d’échantillon d’environ 5 mg.
Une méthode par centrifugation a été modifiée et adaptée afin d’évaluer la valeur de rétention d'eau (WRV) conformément à la norme ISO 23714. Des lots de 0,3 g de faisceaux de fibres ont été immergés dans de l'eau déionisée pendant 5 minutes à 23 °C puis placés dans des tubes pour être centrifugés à 3000 g pendant un temps de 30 minutes et enfin pesés afin de déterminer la masse Mf (en g) des fibres.
Les teneurs en eau en immersion (WRV) ont été déterminées selon l’équation (Eq. 1) après séchage pendant 10 min à 105 °C dans un dessiccateur d’humidité IR Sartorius-MA160 (Grosseron SAS, Coueron, France) et pesée de la masse Ms anhydre des fibres (en g).
$\mathrm{WRV}(\%) \cdot=\frac{M_{f}-M_{S}}{M_{S}} \times 100$ (1)
2.2.2 Caractérisation dimensionnelle
Un dispositif d'analyse dimensionnelle appelé « FDAS » (Fibre Dimensional Analysis System, Diastron Ltd, Hampshire, Royaume-Uni) a été utilisé pour mesurer les dimensions transversales des faisceaux de fibres d’une longueur de 30 mm et ainsi estimer l’aire de section droite qui est généralement évolutive le long des faisceaux de fibres végétales. Cette méthode expérimentale basée sur le balayage laser automatisé de l’objet ainsi que le protocole de préparation des échantillons a été décrite en détail dans une étude précédente [15]. Au cours de la mesure, l’échantillon est mis en translation et en rotation, permettant ainsi de collecter jusqu’à 600 valeurs de diamètre apparent par révolution sur une section, chaque section étant séparée d’un pas longitudinal minimum de 40 µm. La non-circularité des sections transversales des faisceaux de fibres est ainsi mesurée et prise en compte. Pour chaque faisceau de fibres, un modèle elliptique utilisant un traitement statistique des diamètres apparents obtenus pour 42 sections transversales (équiréparties sur une longueur de 30 mm) a été appliqué afin d’estimer leur aire médiane de section transverse dans les différentes conditions hygrothermiques étudiées.
Afin d'analyser l'influence de l'humidité relative sur les dimensions transversales des faisceaux de fibres, l’appareil d'analyse dimensionnelle (FDAS) a été placé dans une chambre climatique fournie par la société ETS (Electro-Tech Systems Inc, Pennsylvanie, USA) pour réguler la température à 23 °C et l’humidité relative à 20, 50 et 73 % HR. Une cellule d'immersion «DSM» (Dynamic Swelling Module, Diastron Ltd., Hampshire, Royaume-Uni) adaptée sur le dispositif FDAS a permis de mesurer les dimensions des faisceaux de fibres en immersion dans de l'eau déionisée à 23 °C. A noter que dans les conditions d’immersion, le dispositif ne permet le suivi dimensionnel des faisceaux de fibres que sur une seule section transverse à la fois. Pour chaque espèce végétale et chaque condition hygro/hydrothermique, 5 à 10 faisceaux de fibres ont été caractérisés en procédant au préalable à une période d'acclimatation de 13 h à 23 °C, à l'HR étudiée (20, 50, 73 %) ou à 50 % d'HR avant immersion.
3.1 Influence des conditions hydro/hygrothermiques sur la teneur en eau
La teneur en eau à l’équilibre des différentes espèces de fibres végétales en fonction du taux d’humidité relative a été quantifiée à partir des courbes d’adsorption obtenues par analyse DVS. Quelle que soit l’espèce végétale considérée, l’allure des courbes est identique et de type sigmoïde (type II) ; elle résulte de la combinaison de différents mécanismes de sorption. Ainsi, les isothermes de sorption peuvent être divisées en trois zones [6]. Dans la première zone, entre 0 et 10 % HR, l’eau est principalement absorbée et liée via des liaisons hydrogènes aux hydroxyles des biopolymères amorphes (pectines, hémicelluloses et cellulose amorphe) présents dans les parois cellulaires et la lamelle mitoyenne, constituant ainsi une première couche mono-moléculaire d’eau sur les sites privilégiés d’adsorption. Dans la seconde zone, entre 10 et 60 % HR, l’eau est absorbée en multicouches au sein des parois cellulaires et du réseau microporeux. Enfin, au-delà de 60 % HR, sont mis en jeu les processus de condensation capillaire qui se traduisent par la formation d’eau libre dans les pores et les lumens jusqu’à saturation en eau des faisceaux de fibres végétales.
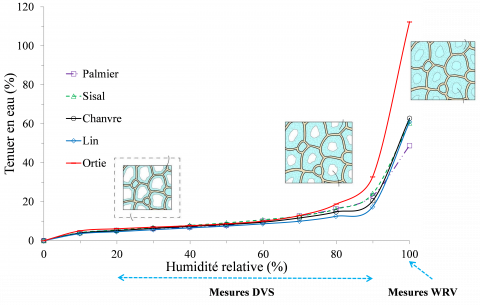
La Figure 2 présente l’évolution de la teneur en eau des faisceaux de fibres végétales en fonction de l’humidité relative (mesures DVS) et en immersion (mesures WRV). Entre 0 et 50 % HR la reprise en eau des différentes espèces végétales est faible et relativement identique, soit en moyenne 5,3 ± 0,4 %. Entre 50 et 73 % HR, on constate une augmentation de la teneur en eau jusqu’à environ 13,7 ± 1,7 % quelle que soit l’espèce végétale considérée. En revanche, au voisinage du point de saturation des parois cellulaires (teneur en eau d’environ 30 %) et dans les conditions d’immersion (formation d’eau libre dans les pores et les lumens), la teneur en eau augmente considérablement et de grandes différences sont observées en fonction des espèces de fibres végétales. Les faisceaux de fibres de palmier et de sisal présentent les plus faibles teneurs en eau en immersion, 48,8 ± 2,0 % et 60,4 ± 1,0 % respectivement, suivis des faisceaux de fibres de lin et de chanvre avec des teneurs en eau respectivement de 60,9 ± 0,7 % et de 62,8 ± 0,7 %. On note que les faisceaux de fibres d’ortie présentent un caractère fortement hygroscopique avec une teneur en eau de 112,2 ± 1,2 %, soit pratiquement deux fois supérieure aux faisceaux de lin et de chanvre.
La composition biochimique contrastée entre les espèces végétales considérées peut expliquer ces différences de sorption d’eau en conditions hygro/hydrothermiques contrôlées. Pejic et al. [19] ont étudié l'influence des biopolymères non-cellulosiques tels que la lignine sur l'absorption d'eau des faisceaux de fibres de chanvre, et ont montré que la diminution de la teneur en lignine tend à augmenter la capacité de rétention d'eau des fibres. Les résultats de cette étude montrent que les faisceaux de fibres de palmier et de sisal qui présentent des teneurs en lignine plus importantes (Tableau 1), sont également ceux qui présentent les plus faibles reprises en eau en immersion. Les faisceaux de fibres libériennes de lin, de chanvre et d’ortie présentent quant à eux des teneurs en lignine plus faibles et des reprises en eau en immersion plus importantes. Une des hypothèses pour expliquer la forte reprise en eau des faisceaux de fibres d’ortie, comparée aux autres fibres libériennes, serait un mauvais rouissage induisant une teneur en pectine élevée (Tableau 1).
La connaissance des teneurs en eau mesurées par DVS et WRV va permettre d'analyser les variations morphologiques et dimensionnelles des faisceaux de fibres végétales dans les différentes conditions hygro/hydrothermiques.
Figure 2. Teneur en eau en phase vapeur (DVS) et en immersion (WRV) pour les cinq espèces végétales à 23 °C
3.2 Etude des variations morphologique et dimensionnelle en fonction de la teneur en eau
3.2.1 Variation morphologique : évolution du facteur de forme de la section transversale
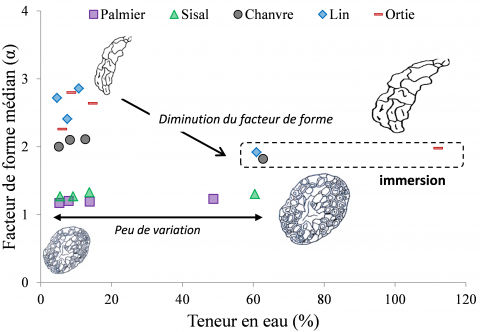
La Figure 3 présente, pour les différentes espèces de fibres végétales, l'évolution du facteur de forme médian (α) de leur section transversale, c’est-à-dire la valeur médiane du rapport diamètre maximum / diamètre minimum mesuré pour chacune des sections du faisceau de fibres considéré, selon les conditions d’humidité. Le facteur de forme est un indicateur morphométrique de la non-circularité de la section transversale des fibres végétales [15], et son évolution est étudiée ici en fonction de la teneur en eau de chaque faisceau de fibres végétales. Dans le cas des faisceaux de fibres de palmier et de sisal, l'humidité relative a peu d'influence sur les facteurs de forme qui restent proches de 1, c'est-à-dire qu’ils conservent des sections de forme quasi-circulaire. En effet, leurs facteurs de forme médians restent similaires, soit environ 1,20 ± 0,06 et 1,29 ± 0,14 respectivement pour le palmier et le sisal, sur toute la gamme de teneur en eau étudiée (soit de 20 % HR à l’immersion). On peut donc conclure que le gonflement de leur section transversale est isotrope.
En revanche, les faisceaux de fibres libériennes présentent des facteurs de forme médians plus élevés et plus disperses, qui sont influencés par la teneur en eau, en particulier en immersion. Le gonflement de ces faisceaux est anisotrope, car il est caractérisé par une augmentation du diamètre minimum très supérieure à celle du diamètre maximum, conduisant à une diminution significative du facteur de forme de section en conditions d’immersion (teneurs en eau supérieures à 60 %). En effet, l’accroissement du diamètre minimum pour les faisceaux de fibres de lin et de chanvre est environ deux fois supérieur à celui du diamètre maximum, et jusqu’à trois fois supérieur pour les faisceaux de fibres d’ortie. Les faisceaux de fibres de chanvre, de lin et d'ortie présentent ainsi des facteurs de forme médians en immersion de 1,82 ± 0,26 ; 1,92 ± 0,42 et 1,98 ± 0,69 respectivement, ce qui implique une augmentation de leur circularité dans ces conditions d’humidité. Au sein d’une tige, les fibres qui poussent en périphérie d’une lacune, entre l’écorce et le xylème, sont regroupées en faisceaux écrasés. Par conséquent, on peut supposer que ce mode de croissance intrusive de ces fibres [20], ainsi que le dessèchement progressif des cellules en fin de croissance, après récolte et coupe des tiges ou des feuilles, entraînent ce phénomène de gonflement anisotrope lors de la réhydratation des fibres élémentaires. Ainsi, plus l'épaisseur des parois cellulaires sera faible, plus l'effet d'aplatissement sera important lors de la dessiccation et plus la diminution du facteur de forme lors de la réhydratation sera importante et la géométrie de la section tendra vers une forme circulaire.
Figure 3. Evolution du facteur de forme médian (α) de la section transversale en fonction de la teneur en eau à 23 °C pour les différents faisceaux de fibres végétales
3.2.2 Variation dimensionnelle : évolution de l’aire médiane de la section transversale et processus de gonflement
L'influence de la teneur en eau sur les variations dimensionnelles des faisceaux de fibres végétales, et en particulier sur le gonflement de leur section transversale, a été analysée. Le gonflement médian des différents faisceaux de fibres végétales à une humidité relative donnée et normalisée par rapport à l’aire médiane de section transversale à 50 % HR, a été reportée pour chaque teneur en eau (soit pour des humidités relatives de 20 % HR jusqu’à l’immersion). On définit le gonflement Gs d’un faisceau de fibre comme suit (Eq. 2) :
$G_{S}(\%)=\frac{S_{i}-S_{o}}{S_{o}} \times 100$ (2)
avec Si l’aire médiane de section transversale du faisceau de fibre considéré pour une humidité donnée (en µm²) et S0 son aire médiane de section transversale à 50 % HR (en µm²).
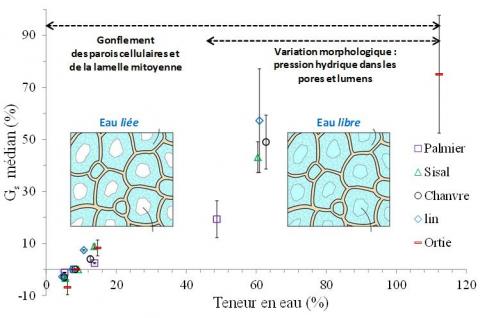
A 20 % HR, soit pour des teneurs en eau inférieures à 5 %, on observe (Figure 4) une faible contraction, allant de 1,2 ± 0,1 % à 3,3 ± 1,0 % pour les faisceaux de fibres de palmier et de sisal, respectivement. Les faisceaux de fibres de lin et de chanvre présentent quant à eux des contractions de 2,8 ± 0,6 % et 3,0 ± 1,8 %, respectivement. Ce phénomène est plus prononcé dans le cas des faisceaux de fibres d'ortie, atteignant 6,9 ± 2,9 %. À 73 % HR, pour des teneurs en eau comprises entre 10,8 % et 14,7 % selon le type de fibres, le gonflement est relativement faible pour toutes les espèces (inférieur à 10 %) avec une faible dispersion. Dans cette gamme d’humidité relative (20 % HR à 73 % HR) correspondant à des teneurs en eau d’environ 5 % à 15 %, le gonflement des faisceaux de fibres végétales est principalement associé à des processus s’opérant au niveau des parois cellulaires et de la lamelle mitoyenne du fait de la sorption d’eau liée par les biopolymères.
Dans les conditions d'immersion, le gonflement médian de la section transversale augmente considérablement et de grandes différences sont observées en fonction des espèces de fibres végétales. On note également une plus grande dispersion sur les valeurs de gonflement. Les faisceaux de fibres de palmier présentent le plus faible gonflement, soit 19,3 ± 7,1 %, suivis par les faisceaux de fibres de sisal avec 43,2 ± 5,9 %. Les faisceaux de fibres de chanvre, de lin et d’ortie montrent des gonflements plus importants, de 49,0 ± 10,4 % ; 57,2 ± 19,9 % et 75,1 ± 22,6 %, respectivement. Un second processus est ici impliqué dans le gonflement des faisceaux de fibres végétales. En effet, la formation d’eau libre en quantité importante dans les lumens et les porosités contribue également au gonflement du faisceau, du fait des variations morphologiques relevées précédemment et induites par la pression hydrique s’exerçant sur les parois cellulaires. Ainsi, le gonflement anisotrope observé en immersion pour les fibres libériennes (lin, chanvre et ortie) participe à l’augmentation de l’aire de section apparente mesurée au FDAS, et donc au gonflement médian mesuré.
Figure 4. Gonflement médian de la section transversale (normalisée par rapport à l’aire de section médiane à 50 % HR) en fonction de la teneur en eau, à 23 °C pour les différents faisceaux de fibres végétales
Le gonflement des faisceaux de fibres végétales est donc gouverné par deux processus (Figure 4) : (i) un gonflement microscopique, à l’échelle des parois cellulaires et de la lamelle mitoyenne, associé à la sorption d’eau liée et, (ii) un gonflement macroscopique associé à la formation d’eau libre dans les pores et lumen (par condensation capillaire ou lors de l’immersion) qui induit une déformation anisotrope des faisceaux de fibres et un accroissement significatif de leur gonflement. Dans le cas des fibres végétales, le point de saturation en eau des parois cellulaires est ainsi difficile à définir car, pour les humidités relatives importantes comprises entre 80 et 99 % HR, le gonflement des parois associé à la sorption d’eau liée et les variations morphologiques associées à la formation d’eau libre par les processus de condensation capillaire sont concomitants. S’agissant des fibres végétales, on parlera donc davantage d’un continuum de processus de gonflement intervenant aux échelles microscopique et macroscopique sans qu’un point de saturation en eau puisse être clairement identifié. Des expérimentations dans une gamme d’humidité relative intermédiaire, c.-à-d. 80 et 99 % HR) seraient néanmoins intéressantes afin de compléter ces données expérimentales. On notera que les dispositifs de mesures utilisés pour cette étude ne permettent cependant pas de travailler dans cette gamme d’humidité du fait des risques de condensation d’eau et d’endommagement des appareils. Une élévation de la température au-delà de 23 °C pourrait permettre d’explorer des humidités relatives plus importantes (potentiellement jusqu’à 90 % HR).
3.3 Détermination des coefficients d’hygro / hydroexpansion surfaciques
L’étude des variations morphologique et dimensionnelle des différents faisceaux de fibres végétales en fonction de leur teneur en eau permet de déterminer leurs coefficients d’expansion hydrique, défini dans cette étude comme le gonflement Gs (%) induit par une variation de 1 % de la teneur en eau Dm (%). Il s’agit donc d’un coefficient d’expansion hydrique surfacique bs. Par ailleurs, nous avons mis en évidence précédemment que le gonflement des faisceaux de fibres végétales résulte de deux phénomènes qui dépendent fortement des conditions hygro/hydrothermiques. Le gonflement des parois cellulaires et de la lamelle mitoyenne associé à la sorption d’eau liée est le principal processus en jeu en conditions hygrothermiques modérées. En conditions hydrothermiques ou dès lors que l’humidité relative est suffisante pour générer la formation d’eau libre (par condensation capillaire), la pression hydrique dans les pores et lumens engendre des variations morphologiques importantes qui participent au gonflement des faisceaux de fibres végétales. Il convient donc de différencier les coefficients d’hygroexpansion surfacique bs,hygro (Eq. 3) et d’hydroexpansion surfacique bs,hydro (Eq. 4), définis comme suit dans cette étude :
$\operatorname{De} 20 \dot{\mathrm{a}} 73 \% \mathrm{HR} \beta_{s, h y g r o}=\frac{G_{s}(\%)}{\Delta m(\%)}$ (3)
En immersion $\quad \beta_{s, h y d r o}=\frac{G_{s}(\%)}{\Delta m(\%)}$ (4)
La Figure 5 présente les coefficients d’hygroexpansion surfaciques bs,hygro pour les différents types de fibres végétales, calculés entre 20 et 73 % HR à 23 °C. Ce coefficient varie de 0,42 à 1,70 selon le type de fibres végétales. A titre comparatif, Le Duigou et al. [13] ont déterminé un coefficient d’hygroexpansion radial de fibres élémentaires de lin (issues de Flax Tape, Lineo®) de 1,14 par des mesures de diamètre effectuées avec un microscope électronique à balayage en environnement contrôlé (23 °C, pression de vapeur 5-10 mbar, 20 % à 98 % HR). A partir du coefficient d’hygroexpansion surfacique obtenu pour le lin dans cette étude, on peut déduire un coefficient d’hygroexpansion radial équivalent ($\left(\beta_{r}=\sqrt{1+\beta_{s}}-1\right)$) dont la valeur est 0,63. Cette valeur obtenue pour un faisceau de fibres de lin s’avère être inférieure à celle obtenue par Le Duigou et al. [13] pour une fibre élémentaire de lin. Joffre et al. [21] ont déterminé un coefficient d’hygroexpansion local (dans le plan transverse aux microfibrilles de cellulose) de fibres de bois (épicéa, Picea abies) de 0,45 en effectuant des mesures par micro-tomographie aux rayons X (Synchrotron, Grenoble) et une identification par méthode numérique (23 °C, 47 % à 80 % HR). Bien que ces coefficients d’hygroexpansion soient du même ordre de grandeur, ces différents résultats illustrent l’importance de la méthode de mesure, des dimensions et de la nature de l’objet considérées. En effet, selon l’échelle (micro / nano) et le type de fibres végétales étudiés, les variations induites de microstructure et de composition biochimique auront une influence sur la valeur du coefficient d’hygroexpansion. On notera par ailleurs la forte anisotropie des microstructures rencontrées dans les fibres végétales qui engendre des écarts très importants entre les coefficients d’hygroexpansion transversal et longitudinal, ce dernier étant très faible en particulier pour les fibres végétales possédant un faible MFA (ex : 0,037 pour les fibres de bois dans l’étude de Joffre et al. [21]).
Figure 5. Détermination des coefficients d’hygroexpansion surfaciques bs,hygro des différents faisceaux de fibres végétales mesurés entre 20 % et 73 % HR à 23 °C
De la même façon, les coefficients d’hydroexpansion surfaciques bs,hydro ont été déterminés pour les différents types de faisceaux de fibres végétales et sont reportés dans le Tableau 2. De façon générale, on constate que ces coefficients sont inférieurs aux coefficients d’hygroexpansion surfaciques bs,hygro, en particulier pour les faisceaux de fibres qui présentent de fortes reprises en eau et/ou variations morphologiques en immersion, c.-à-d. le lin et l’ortie (Figures 3 et 4). Ceci indique que la sorption d’eau liée au sein des parois cellulaires et de la lamelle mitoyenne joue un rôle prépondérant sur le gonflement des faisceaux de fibres végétales. On remarquera aussi que, pour des espèces phylogénétiquement éloignées telles que le palmier et le chanvre, les valeurs des coefficients bs, hygro et bs, hydro obtenues sont similaires.
Tableau 2. Coefficients d’hygroexpansion bs,hygro et d’hydroexpansion bs,hydro surfaciques à 23 °C des différents faisceaux de fibres végétales
|
Espèce végétale |
bs, hygro |
bs, hydro |
|
Palmier |
0,42 |
0,47 |
|
Sisal |
1,49 |
0,84 |
|
Chanvre |
0,92 |
0,90 |
|
Lin |
1,67 |
1,07 |
|
Ortie |
1,70 |
0,72 |
Le Tableau 3 propose une corrélation (Pearson, r) entre le coefficient d’hygroexpansion surfacique bs,hygro et les caractéristiques microstructurales des parois cellulaires des fibres végétales telles que leur composition biochimique et leur MFA. Cette analyse met en évidence une forte corrélation du coefficient bs,hygro avec la teneur massique en lignine et le MFA avec un coefficient r de -0,76 et -0,77, respectivement. Ainsi, les plus fortes teneurs en lignine et les MFA élevés rencontrés notamment dans les faisceaux de fibres de palmier tendent à limiter la capacité de rétention d'eau, à réduire le gonflement transverse (Figure 4), et donc à diminuer le coefficient d’hygroexpansion surfacique (0,42). A l’inverse la faible teneur en lignine et le faible MFA rencontrés dans les faisceaux de fibres de lin sont corrélés avec un plus fort coefficient d’hygroexpansion surfacique (1,67). Des approches statistiques de type analyses multivariées sur des ensembles de données plus étendus en termes de composition biochimique des parois cellulaires, de MFA et intégrant d’autres descripteurs tels que la composition/structure des lamelles mitoyennes, la taille des lumens… pourraient permettre de mieux comprendre la contribution de ces différents paramètres structuraux au gonflement des faisceaux de fibres végétales.
Tableau 3. Corrélation (coefficient de Pearson, r) entre les coefficients d’hygroexpansion surfaciques bs,hygro (20 % - 73 % HR ; 23 °C) et les compositions biochimiques et MFA des différents faisceaux de fibres végétales
|
Espèce végétale |
bs, hygro |
Cellulose (%) |
Hémicellulose (%) |
Autres (pectines…) (%) |
Lignine (%) |
MFA (°) |
|
Palmier |
0,42 |
45,1 ± 3,4 |
27,7 ± 4,9 |
10 |
16,2 ± 0,3 |
43,5 |
|
Sisal |
1,49 |
60,1 ± 1,2 |
23,3 ± 2,1 |
8 |
9,3 ± 0,1 |
20 |
|
Chanvre |
0,92 |
68,8 ± 1,3 |
9,8 ± 1,8 |
17 |
4,7 ± 0,3 |
8,5 |
|
Lin |
1,67 |
64,9 ± 2,9 |
9,9 ± 3,9 |
21 |
3,7 ± 0,3 |
8 |
|
Ortie |
1,70 |
51,7 ± 0,9 |
9,9 ± 1,2 |
34 |
4,6 ±0,2 |
7,5 |
|
Coef. Pearson (r) |
0,36 |
-0,58 |
0,56 |
-0,76 |
-0,77 |
|
L’effet des conditions d’humidité sur la teneur en eau et les variations dimensionnelles de différents faisceaux de fibres végétales (palmier, sisal, chanvre, lin, ortie) aux caractéristiques biochimiques et structurelles contrastées a été étudié par des mesures de DVS et WRV et une technique de mesure de la section transversale des faisceaux par balayage laser automatisé conduite dans une chambre climatique et dans une cellule d'immersion. Les résultats ont mis en évidence des processus de reprise en eau et de gonflement différenciés selon les espèces végétales et les conditions hygro/hydrothermiques : (i) un gonflement microscopique, à l’échelle des parois cellulaires et de la lamelle mitoyenne, associé à la sorption d’eau liée et, (ii) un gonflement macroscopique associé à la formation d’eau libre dans les pores et lumens (par condensation capillaire ou lors de l’immersion) qui induit une déformation anisotrope des faisceaux de fibres et contribue à leur gonflement. Ces résultats ont permis la détermination des coefficients d’hygro/hydroexpansion surfaciques des différentes fibres végétales étudiées variant de 0,42 à 1,70. Une corrélation du coefficient d’hygroexpansion avec les caractéristiques structurales des parois cellulaires des fibres végétales (composition biochimique et MFA) est proposée, mettant ainsi en évidence une dépendance à la teneur en lignine et au MFA. Ces résultats ouvrent des perspectives intéressantes pour la modélisation prédictive du comportement mécanique «en service» des biocomposites qui prendrait en compte le gonflement en humidité des fibres végétales en lien avec leurs caractéristiques structurales.
William Garat remercie Montpellier SupAgro pour le financement de ses travaux de doctorat. Les auteurs remercient Arnaud Day (Fibres Recherche Développement, Troyes, France) pour la fourniture des fibres d’ortie, et Yann Leray et Steve Bucknell (Diastron Ltd., Hampshire, Grande-Bretagne) pour leur support technique sur les appareils FDAS et DSM.
|
aw |
activité de l’eau |
|
P |
pression de vapeur d’eau à la surface du substrat considéré, Pa |
|
P0 |
pression de vapeur saturante de l’eau pure, Pa |
|
Gs |
gonflement d’un faisceau de fibre, % |
|
Mf |
masse des fibres après centrifugation, g |
|
Ms |
masse des fibres anhydre, g |
|
Si |
aire médiane de section transversale pour une humidité donnée, µm² |
|
S0 |
aire médiane de section transversale à 50 % HR, µm² |
|
Symboles grecs |
|
|
α |
facteur de forme médian |
|
△m |
teneur en eau, % |
|
bs |
coefficient d’expansion hydrique surfacique |
|
bs,hygro |
coefficient d’hygroexpansion surfacique |
|
bs,hydro |
coefficient d’hydroexpansion surfacique |
|
br |
coefficient d’hygroexpansion radial équivalent |
[1] Placet, V. (2009). Influence de traitements hygrothermiques sur les propriétés mécaniques de composites à fibres végétales. Actes des JNC, 16: 1-10. Toulouse. https://hal.archives-ouvertes.fr/hal-00388784
[2] Placet, V., Cisse, O., Boubakar, M.L. (2012). Influence of environmental relative humidity on the tensile and rotational behaviour of hemp fibres. Journal of Materials Science, 47(7): 3435-3446. https://doi.org/10.1007/s10853-011-6191-3
[3] Zelinka, S.L., Glass, S.V., Jakes, J.E., Stone, D.S. (2016). A solution thermodynamics definition of the fiber saturation point and the derivation of a wood-water phase (state) diagram. Wood Science and Technology, 50: 443-462. https://doi.org/10.1007/s00226-015-0788-7
[4] Bourmaud, A., Dhakal, H., Habrant, A., Padovani J., Siniscalco, D., Ramage, MH., Beaugrand, J., Shah, D.U. (2017). Exploring the potential of waste leaf sheath date palm fibres for composite reinforcement through a structural and mechanical analysis. Composites Part A: Applied Science and Manufacturing, 103: 292-303. https://doi.org/10.1016/j.compositesa.2017.10.017
[5] Guicheret-Retel, V., Cisse, O., Placet V., Beaugrand, J., Pernes, M., Boubakar, M.L. (2015). Creep behaviour of single hemp fibres. Part II: Influence of loading level, moisture content and moisture variation. Journal of Materials Science, 50: 2061-2072. https://doi.org/10.1007/s10853-014-8768-0
[6] Hill, C.A.S., Norton, A., Newman, G. (2009). The water vapor sorption behavior of natural fibers. Journal of Applied Polymer Science, 112: 1524-1537. https://doi.org/10.1002/app.29725
[7] ISO 23714:2014 (2014). Pulps - Determination of water retention value (WRV), pp. 1-8. https://www.iso.org/standard/60371.html
[8] «Wood Movement - Workshop Companion», http://www.workshopcompanion.com/, consulté le 05 octobre 2018.
[9] Burgert, I., Eder, M., Gierlinger, N., Fratzl, P. (2007). Tensile and compressive stresses in tracheids are induced by swelling based on geometrical constraints of the wood cell. Planta, 226: 981-987. https://doi.org/10.1007/s00425-007-0544-9
[10] Lu, Y., Pignatello, J.J. (2002). Demonstration of the “conditioning effect” in soil organic matter in support of a pore deformation mechanism for sorption hysteresis. Environmental Science & Technology, 36: 4553-4561. https://doi.org/10.1021/es020554x
[11] Salmén, L., Larsson, P.A. (2018). On the origin of sorption hysteresis in cellulosic materials. Carbohydrate Polymers, 182: 15-20. https://doi.org/10.1016/j.carbpol.2017.11.005
[12] Karmaker, A.C., Hinrichsen, G. (1994). Effect of water uptake on some physical properties of jute fibres. The Journal of The Textile Institute, 85: 288-290. https://doi.org/10.1080/00405009408659028
[13] Le Duigou, A., Merotte, J., Bourmaud, A., Davies, P., Belhouli, K., Baley C. (2017). Hygroscopic expansion: A key point to describe natural fibre/polymer matrix interface bond strength. Composites Science and Technology, 151: 228-233. https://doi.org/10.1016/j.compscitech.2017.08.028
[14] Regazzi, A., Corn, S., Ienny, P., Bénézet, J.C., Bergeret, A. (2016). Reversible and irreversible changes in physical and mechanical properties of biocomposites during hydrothermal aging. Industrial Crops and Products, 84: 358-365. https://doi.org/10.1016/j.indcrop.2016.01.052
[15] Garat, W., Corn, S., Le Moigne, N., Beaugrand, J., Bergeret, A. (2018). Analysis of the morphometric variations in natural fibres by automated laser scanning: towards an efficient and reliable assessment of the cross-sectional area. Composites Part A: Applied Science and Manufacturing, 108: 114-123. https://doi.org/10.1016/j.compositesa.2018.02.018
[16] Baley, C. (2013). Fibres naturelles de renfort pour matériaux composites. Techniques de L’Ingénieur, AM5130 v2, pp. 1-17.
[17] Bodros, E., Baley, C. (2008). Study of the tensile properties of stinging nettle fibres (Urtica dioica). Materials Letters, 62(14): 2143-2145. https://doi.org/10.1016/j.matlet.2007.11.034
[18] Müssig, J., Fischer, H., Graupner, N., Drieling, A. (2010). Testing methods for measuring physical and mechanical fibre properties (plant and animal fibres). In: Müssig, J. (ed) Industrial applications of natural fibres: structure, properties and technical applications. Wiley, West Sussex, UK, pp. 269-309. https://doi.org/10.1002/9780470660324
[19] Pejic, B.M., Kostic, M.M., Skundric, P.D., Praskalo, J.Z. (2008). The effects of hemicelluloses and lignin removal on water uptake behavior of hemp fibers. Bioresource Technology, 99(15): 7152-7159. https://doi.org/10.1016/j.biortech.2007.12.073
[20] Chernova, T.E., Mikshina, P.V., Salnikov, V.V., Ibragimova, N.N., Sautkina, O.V., Gorshkova, TA. (2018). Development of distinct cell wall layers both in primary and secondary phloem fibers of hemp (Cannabis sativa L.). Industrial Crops and Products, 117: 97-109. https://doi.org/10.1016/j.indcrop.2018.02.082
[21] Joffre, T., Isaksson, P., Dumont, P.J.J., Rolland du Roscoat, S., Sticko, S., Orgeas, L., Gamstedt, K. (2016). A method to measure moisture induced swelling properties of a single wood cell. Experimental Mechanics, 56(5): 723-733. https://doi.org/10.1007/s11340-015-0119-9