Impact of Field Retting on the Hemp Fibres Structure
Impact du Rouissage en Champ sur la Structure des Fibres de Chanvre

Brahim Mazian* | Anne Bergeret | Jean-Charles Benezet | Sandrine Bayle | Luc Malhautier
OPEN ACCESS
In the hemp industry, retting is considered as an upstream processing treatment that facilitates the decortication of fibres in the central woody part of the stem. This treatment is currently carried out empirically in the fields, and leads to variability in the color, (micro)structure and intrinsic properties (thermal properties) of hemp fibers. The present study aims to compare unretted and retted hemp fibres harvested at the end of flowering. The results show that retting induces a change in the color of fibres from light green to dark gray due to the activity of the microbial community at the stem surface. A separation of the fibre bundles into elementary fibres occurs with the degradation of the cementing compounds present in the middle lamella. The cellulose and its crystallinity degree increased from 68 to 75 % and 85 to 89 %, respectively, with a removal of non-cellulosic materials, which leads to an improvement of the thermal stability for the retted fibres.
RÉSUMÉ:
Dans l'industrie du chanvre, le rouissage est considéré comme un premier traitement de transformation qui permet de faciliter la décortication des fibres de la partie centrale de la tige. Ce traitement est actuellement réalisé de façon empirique dans les champs, ce qui entraîne une variabilité de l’aspect, de la (micro)structure et des propriétés thermiques des fibres de chanvre. La présente étude vise à comparer des fibres de chanvre récoltées en fin de floraison, non-rouies et rouies pendant 9 semaines en champ. Les résultats révèlent que le rouissage induit un changement de la couleur des fibres du vert clair au gris foncé en raison d’un développement de la communauté microbienne (champignons et bactéries) à la surface de la tige. Une séparation des faisceaux de fibres en fibres élémentaires se produit avec la dégradation des « ciments » présents dans la lamelle mitoyenne. Le taux de cellulose et son degré de cristallinité augmentent de 68 à 75 % et de 85 à 89 % respectivement, avec une diminution de la matière non-cellulosique, ce qui conduit à une amélioration de la stabilité thermique des fibres rouies.
retting, hemp fibres, chemical composition, thermal stability, crystallinity
Mots-clés :
rouissage, fibres de chanvre, composition chimique, stabilité thermique, cristallinité
La récente prise en compte des facteurs environnementaux et économiques dans l’élaboration des matériaux contraint les industriels à développer de nouveaux matériaux ayant un impact moindre sur l’environnement et pour lesquels les coûts des matières premières et de fabrication sont modérés. L’utilisation de ressources renouvelables, comme les fibres végétales en tant que renforts dans les matériaux composites dits biosourcés (biocomposites) permettrait d’intégrer ces contraintes environnementales et économiques [1]. En plus de leur disponibilité, ces fibres présentent également des atouts liés directement à leurs propriétés intrinsèques comme la densité [2], la biodégradabilité [3], les propriétés mécaniques [4-5], et indirectement, le coût de fabrication des matériaux biosourcés. Un certain nombre de produits sont actuellement commercialisés sur le marché, notamment dans le domaine de l’automobile (panneaux de porte, panneaux de garniture de toit, doublures de démarrage, banquettes arrières, etc.) et d’autres secteurs industriels comme l’industrie nautique ou la construction. Néanmoins, la variabilité des propriétés physicochimiques et de la stabilité thermique des fibres végétales a été constatée dans différentes applications. De plus, le niveau d’adhérence entre la fibre végétale et la matrice ainsi qu’une durée de vie limitée dans certaines conditions d’utilisation constituent des freins majeurs à une utilisation plus large de ces fibres. Une des voies pour pallier à ces verrous est d’être en mesure de mieux contrôler l’ensemble des étapes de transformation des fibres végétales depuis le champ jusqu’à l’obtention de la fibre technique.
La fibre de chanvre a fait l’objet de nombreux travaux de recherche et possède des propriétés compatibles avec une utilisation dans le cadre de la fabrication de matériaux biosourcés. Cette fibre, comme la fibre de lin, subit un traitement préliminaire de rouissage (première étape de transformation dans les champs) qui consiste à couper et mettre les tiges de chanvre en contact avec le sol, où elles sont colonisées par des microorganismes (champignons et bactéries) produisant notamment des enzymes pectinolytiques. Ce développement microbien entraîne l’élimination de certains composés cimentiques qui assurent le lien entre les faisceaux de fibres [6]. Ce traitement de rouissage est réalisé à l’heure actuelle de manière empirique et conduit à une variabilité des caractéristiques des fibres de chanvre en termes de couleur, de microstructure, de composition chimique et de propriétés thermiques et mécaniques.
L’objectif de ce travail est donc de mieux comprendre les mécanismes se déroulant pendant l’étape de rouissage des fibres de chanvre. Une approche multi-échelle et systémique est mise en place afin d’identifier des indicateurs pertinents du rouissage permettant à la fois aux exploitants agricoles, aux transformateurs de fibres végétales et aux applicateurs des matériaux une meilleure maîtrise des performances des fibres de chanvre selon l’application finale visée.
2.1 Matériaux
Le chanvre (Cannabis Sativa L., variété Santhica 27) a été cultivé dans le sud de la France autour du Mas La Valus (GPS : N 44.130673°, E 4.315895°) par des agriculteurs membres de l’Association Chanvre Gardois CIVAM (Le Bouquet, France).
La densité de semis est de 30 kg/ha. Les graines de chanvre ont été semées le 6 mai 2016. Les tiges de chanvre ont été coupées manuellement à différentes périodes : début, fin de floraison et période de semence à l'aide d'une cisaille. Elles ont ensuite été rouies en champ par contact direct avec le sol. Les tiges ont été collectées après différents temps de rouissage entre 0 et 9 semaines. Les tiges ont été retournées régulièrement (une fois par semaine) pour que le rouissage soit homogène. Dans cet article, ne seront présentés que les résultats relatifs aux tiges coupées à la fin de la floraison (tiges non rouies, NR) et collectées après 9 semaines de rouissage (tiges rouies, R).
2.2 Aspect visuel
Le changement de la couleur des tiges et des fibres après un rouissage de 9 semaines a été observé visuellement.
2.3 Observations microscopiques
Le microscope électronique à balayage environnemental (MEBE) Quanta FEG 200 (FEI, Merignac, France) a été utilisé pour observer la surface et la structure des tiges. Avant les observations, les sections transversales des tiges ont été découpées manuellement puis enduites d'une résine époxy (résine Geofix® (ESCIL, Chassieu, France)) et enfin polies à l'aide de différentes feuilles de polissages fins (600, 2400 et 4000 grains) pour obtenir des échantillons parfaitement lisses. Des images microscopiques ont été enregistrées afin de réaliser des analyses qualitatives.
2.4 Extraction et amplification de l'ADN
L’extraction d’ADN des tiges de chanvre a été réalisée en se basant sur la méthode utilisé par [7]. Trois grammes de tiges ont été introduits dans un volume de 15 mL de solution saline (NaCl 0.9 %). Les bactéries sont désorbées par agitation du matériau à 30 Hz pendant 10 min (Vibro-shaker MM200, Retsch, Haan, Allemagne). La phase liquide est ensuite centrifugée à 700 rpm pendant 20 min pour éliminer les particules les plus grossières. Le surnageant est récupéré puis centrifugé à 9000 rpm pendant 30 min à 5 °C. Le culot cellulaire a été récupéré et l'ADN extrait au moyen d’un kit FastDNA® SPIN Kit for Soil (MP Biomedicals, Irvine, CA, USA) selon le protocole donné par le fournisseur. L’ADN génomique est quantifié par absorbance à 535 nm après une coloration au PicoGreen (Quant-IT ds DNA HS reagent, Invitrogen, OR, USA). Les densités de peuplement des communautés bactériennes et fongiques ont été déterminées par PCR quantitative (qPCR) en ciblant les gènes de l’ADNr16S et de l’ADNr18S respectivement. Le protocole permettant de cibler ces deux populations est détaillé ci-dessous.
Les amorces permettant de cibler la communauté bactérienne sont BAC338F (5'-ACTCCTACGGGAGGCAG-3'), BAC805R (5'-GACTACCAGGGTATCTAATCC 3') et la sonde BAC516F (FAM-TGCCA GCAGC CGCGG TAATA C-TAM). La qPCR a été effectuée au moyen du kit Promega GoTaq Probe qPCR Master mix, en thermocycleur Corbett Research Rotorgene 6000 (Qiagen, Hilden, Allemagne) selon le programme suivant : activation initiale à 95 °C pendant 4 min suivie de 50 cycles de : dénaturation à 95 °C pendant 15 s et d’hybridation-élongation à 60 °C pendant 60 s.
Les amorces permettant de cibler la communauté fongique sont nu-SSU-0817-59-5’ (5'-TTAGCATGGAATAATRRAATAGGA-3') et nu-SSU-1196-39-3’ (5'-TCTGGACCTGGTGAGTTTC-3'). Le kit Hot firepol EvaGreen (Solis BioDyne, Tartu, Estonie), en thermocycleur Corbett Research Rotorgene 6000 (Qiagen, Hilden, Allemagne) a été utilisé pour réaliser les quantifications selon le programme suivant : activation initiale à 95 °C pendant 15 min suivie de 50 cycles : dénaturation à 95 °C pendant 18 s, hybridation à 60 °C pendant 60 s et extension à 72 °C pendant 20 s. Les courbes de fusion des amplicons ont été obtenues à partir de la rampe de température suivante : de 65 à 95 °C à 0,5 °C/s.
2.5 Analyse chimique
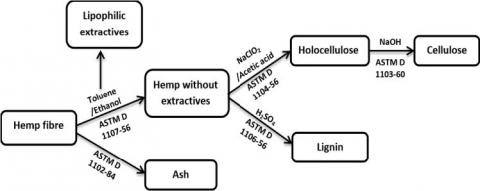
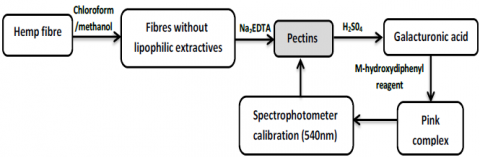
Les analyses biochimiques des fibres de chanvre ont été réalisées en se basant sur deux méthodes [8]. La cellulose, la lignine et les extraits lipophiles ont été quantifiés selon les méthodes d'essai ASTM (Figure 1) qui sont basées sur des extractions successives par solvant. La teneur en pectines a été déterminée par spectrophotométrie (Figure 2).
Figure 1. Analyse de la composition chimique des fibres de chanvre par extraction par solvants [8]
Figure 2. Méthode utilisée pour quantifier la teneur en pectines des fibres de chanvre [8]
2.6 Diffraction des rayons X
La microstructure des fibres de chanvre a été analysée à l’aide d’un diffractomètre de rayons X (XRD, AXS D8 Advance Bruker) équipé d'un rayonnement Cu-Ka (λ = 1,54 Å). Les mesures ont été effectuées sur des fibres coupées et compressées (disques de 25 mm de diamètre et de 2 mm d'épaisseur). Les diffractogrammes ont été enregistrés dans une plage d'angle 2θ de 5° à 70° avec une vitesse de balayage de 0,01°/s. L'indice d'ordre cristallin (IC) a été déterminé à partir de diffractogrammes de rayons X en utilisant la méthode empirique de Segal [9].
2.7 Analyse thermogravimétrique
Les analyses thermogravimétriques des fibres de chanvre non-rouies (NR) et rouies (R) ont été réalisées en utilisant un système d'analyse thermique Perkin-Elmer Pyris-1. Des échantillons d'environ 8 mg de fibres ont été chauffés sous azote de 30 °C à 700 °C à 10 °C/min. La perte de masse a été enregistrée en fonction de la température.
3.1 Evolution de la couleur et des densités microbiennes des tiges de chanvre sur les tiges rouies et non rouies
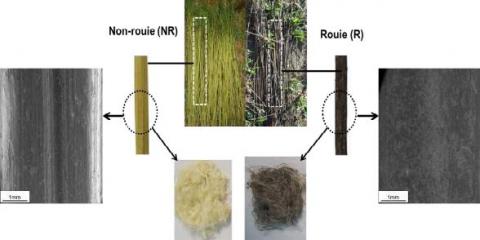
Une observation visuelle des échantillons prélevés après la récolte (fibres NR) et après 9 semaines de rouissage (fibres R) a été réalisée (Figure 3). La couleur varie du vert pâle avant rouissage au gris foncé après rouissage. Ce changement de couleur est lié à l’activité de communautés microbiennes (champignons et bactéries [10]) à la surface de la tige pendant le rouissage en champ. L'observation au MEBE a montré que les hyphes fongiques et bactéries ont colonisé la surface de la tige de chanvre après rouissage. Cela a également été rapporté par Akin et al., 2000 et Liu et al., 2017 [11-12].
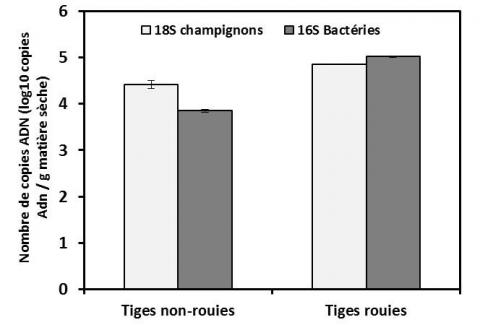
Afin d’associer ces observations qualitatives à une analyse quantitative, les densités de peuplement des communautés bactérienne et fongique à la surface de tiges rouies et non rouies ont été déterminées par qPCR (Figure 4). Une augmentation des densités de peuplement de ces deux communautés est observée. Cependant, une étude plus approfondie de la dynamique temporelle de la structure des communautés microbiennes totales et fonctionnelles est nécessaire pour mieux comprendre les mécanismes biologiques qui se produisent au cours du rouissage en champ et identifier les populations directement impliquées dans la dégradation des composés pectiques.
Figure 3. Photographies de changement de couleur pendant le rouissage et images au MEBE de la surface des tiges de chanvre non rouies (NR) et rouies (R)
Figure 4. Densités de peuplement des populations bactériennes et fongiques à la surface des tiges de chanvre non rouies et rouies
3.2 Evolution de la morphologie des tiges de chanvre au cours du rou
Les images MEBE de sections transversales des tiges de chanvre sont présentées sur la Figure 5. Ces images révèlent l'impact du rouissage sur la morphologie des fibres de chanvre. La structure générale de la tige de chanvre a déjà été décrite par plusieurs auteurs [5, 13]. Les parties principales de la tige de chanvre sont le bois au centre de la tige (xylème) et le cortex qui contient les faisceaux de fibres (incluant les fibres primaires et secondaires), l'épiderme et le parenchyme cortical. Ces deux parties sont liées par le cambium. Les différentes parties de la tige non rouies apparaissent comme intactes et bien organisées sous la forme d’un faisceau (Figure 5A). En revanche, lorsque les tiges sont rouies, la structure des tiges est modifiée. La couche d'épiderme et le cambium ont disparu, les composés présents entre les faisceaux de fibres ont été éliminés, à l’origine de la séparation des faisceaux de fibres en faisceaux de plus petite taille et en fibres élémentaires. Après rouissage, le lumen des fibres élémentaires est observé de manière distincte. Ces changements de la morphologie des fibres de chanvre au cours du processus de rouissage en champ sont dus à l'action des microorganismes qui recouvrent progressivement la surface des tiges. L'activité microbienne pendant le rouissage permettrait l'élimination de la substance intercellulaire, principalement la dégradation des pectines et des extraits lipophiles, aboutissant à une séparation des faisceaux de fibres du bois central et de l'épiderme. Ce résultat confirme l'impact positif du rouissage sur la séparation des faisceaux de fibres en fibres élémentaires.
Figure 5. Images MEBE de coupes transversales de tiges de chanvre A) non rouies (NR) et B) rouies (R). E : épiderme ; CFB : couche des faisceaux de fibres primaires ; CFS : couche des faisceaux de fibres secondaires ; L : lumen ; FB : fibres primaires (élémentaires) ; FS : fibres secondaires (élémentaires) ; EV : espaces vides ; X : xylème
3.3 Evolution de la composition chimique des fibres de chanvre au cours du rouissage
L'évolution de la composition chimique des fibres de chanvre est illustrée sur la Figure 6. La fraction de cellulose augmente significativement de 68 % à 75 % après rouissage, tandis que les fractions de pectines et les hémicelluloses diminuent de 7 % à 4 %, et de 16 % à 15 % respectivement. Ces variations pendant le rouissage ont été rapportées par différents auteurs. Dans le cadre du rouissage enzymatique, Nykter et al., 2008 [14] ont utilisé la même méthode d'extraction et ont constaté le même phénomène. La dégradation des pectines est également en accord avec les données rapportées par Meijer et al., 1995 et Musialak et al., 2008 [15-16] lors du rouissage en champ. D’autres auteurs [5] ont observé une augmentation de la teneur en cellulose au début du traitement du rouissage en champ avec une diminution des composants non cellulosiques (pectines et hémicelluloses). La Figure 6 montre également que les minéraux (cendres) et les extraits lipophiles ont été éliminés pendant le rouissage. En revanche, la teneur en lignine augmente de manière surprenante de 4 % à 9 %. Cette augmentation de la teneur en lignine après le rouissage a également été observée par Liu et al., 2015a et Placet et al., 2017 [5, 17], qui relient cet accroissement à la vitesse de dégradation de la lignine qui est plus lente que celle des autres composants (extraits lipophiles, cendres et carbohydrates) qui sont éliminés dans la même période. Placet et al., 2017 [17] ont suggéré une autre hypothèse en relation avec la méthode de Klason utilisée pour déterminer la teneur en lignine. En effet, par cette méthode, d'autres composants phénoliques ou protéiques qui évoluent pendant le rouissage sont probablement co-analysés avec la lignine.
Figure 6. Composition chimique de fibres de chanvre non rouies (NR) et rouies (R)
Ces résultats semblent indiquer que le traitement du rouissage en champ affecte la composition chimique des fibres de chanvre grâce à l'activité métabolique des microorganismes [10, 12].
3.4 Evolution de la cristallinité des fibres de chanvre au cours du rouissage
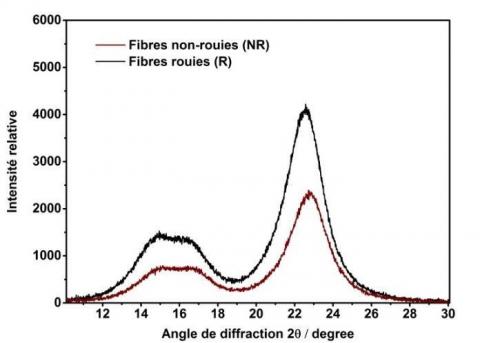
L'influence du rouissage en champ sur l'organisation de la cellulose des fibres a été examinée en utilisant la diffraction des rayons X. Les diffractogrammes sont présentés à la Figure 7. Les échantillons présentent trois pics bien définis et situés à des angles de diffraction 2θ de 14,8°, 16,2° et 22,6°, qui correspondent respectivement aux plans cristallographiques de la cellulose du type I : (101), (101̅) et (002). Lorsque les fibres sont rouies (R), l'intensité des pics cristallins augmente et les pics autour de 14-16° sont plus prononcés, tandis que pour les fibres non rouies (NR), ces pics sont peu différenciés (un seul pic). Cette première indication montre que la cellulose cristalline est élevée pour les fibres rouies et que la présence de la matière amorphe des fibres non rouies ne permet pas de différencier la cellulose cristalline de la matière amorphe dans ces dernières.
Figure 7. Diffractogrammes de fibres de chanvre non rouies (NR) et rouies (R)
Afin de quantifier ce changement, l'indice d'ordre cristallin (IC) a été déterminé. Une augmentation de 85 % à 89 % au cours du rouissage a été observée. Cette observation peut être liée à la dégradation de composés non cellulosiques comme mentionné précédemment dans les analyses chimiques. En effet, la présence d'une grande quantité de composants amorphes dans les fibres de chanvre non rouies, tels que les hémicelluloses, la lignine et les pectines, entre les microfibrilles de cellulose provoquent des zones désorientées qui peuvent influencer leur cristallinité.
3.5 Evolution de la stabilité thermique des fibres de chanvre au cours du rouissage
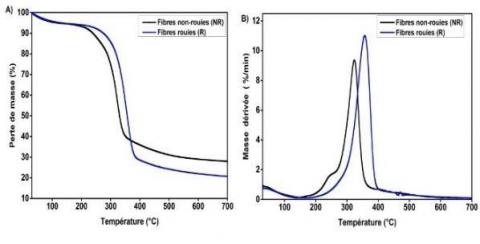
Les analyses thermogravimétriques (ATG) ont été réalisées afin d'évaluer l’évolution de la stabilité thermique des fibres de chanvre au cours du rouissage en champ (Figure 8). Comme déjà décrit dans la littérature [17-18], la perte de masse des fibres de chanvre se déroule en trois étapes, (i) la première à environ 30-100 °C qui est liée à l'évaporation de l’eau contenue dans les fibres, (ii) la seconde aux alentours de 230260 °C qui correspond à la décomposition de la matière non cellulosique (pectines et hémicelluloses), (iii) et la troisième à approximativement 335 °C qui est attribuée à la décomposition du composant majeur des fibres, la cellulose. Une amélioration de la stabilité thermique des fibres de chanvre de 323 °C à 357 °C après rouissage est observée. Par ailleurs, le pic correspondant à la dégradation des composés non cellulosiques (230-260 °C) disparait après rouissage. Cette observation s'explique vraisemblablement par l'élimination de la matière non cellulosique pendant le rouissage, ce qui conduit à une augmentation de la fraction relative de la cellulose comme l’ont montré les analyses chimiques et la diffraction des rayons X.
Figure 8. Perte de masse TG (A) et sa dérivée DTG (B) pour les fibres de chanvre non rouies (NR) et rouies (R)
Cette étude a montré qu’un traitement de rouissage en champ de tiges de chanvre récoltées en fin de floraison pendant 9 semaines conduit à :
(1).Un changement de la couleur des fibres et des tiges du vert pâle au gris foncé, dû à la colonisation de la surface de la tige par des microorganismes (champignons et bactéries) et fort probablement à leur activité métabolique.
(2).Une séparation des faisceaux de fibres en fibres élémentaires en raison de la dégradation du parenchyme cortical, de la lamelle mitoyenne et d'autres composés ciments (pectines, cires).
(3).Une dégradation de la matière non cellulosique avec une augmentation de la fraction de cellulose de 68 % à 75 %, et de son degré de cristallinité de 85 % à 89 %.
(4).Une amélioration de la stabilité thermique des fibres de 35 °C, due à la dégradation de la matière non cellulosique et à l’augmentation de la fraction cellulosique qui nécessite une température de décomposition élevée.
Dans l’objectif de fournir aux différents acteurs de la chaîne de valeur des fibres végétales (exploitants agricoles, transformateurs de fibres, producteurs de matériaux, etc.) des indicateurs simples de suivi du rouissage pour une optimisation des propriétés d’usage des matériaux incorporant ces fibres, il est nécessaire de compléter ces travaux par une meilleure connaissance des communautés microbiennes impliquées dans ce procédé et d’associerl’ensemble de ces données aux propriétés mécaniques des fibres et des matériaux composites intégrant ces fibres végétales. L’influence de la période de récolte des tiges et des conditions climatiques sur le rouissage sont également deux paramètres clés à prendre en compte pour une meilleure maîtrise de ce procédé biologique. Enfin, d’autres critères (émission de composés volatils lors du rouissage par exemple), rarement pris en compte jusqu’à présent, pourraient contribuer à la définition de ces indicateurs.
Les auteurs souhaitent remercier l'association Chanvre Gardois CIVAM (Le Bouquet, France) pour avoir mis à notre disposition une parcelle de terrain pour réaliser cette étude. Les auteurs remercient également Mme Janick Rocher, personnel technique mis à disposition par l’INRA, pour les analyses de PCR quantitative.
[1] Joshi, S.V., Drzal, L.T., Mohanty, A.K., Arora, S. (2004). Are natural fiber composites environmentally superior to glass fiber reinforced composites? Composites Part A: Applied Science and Manufacturing, 35: 371-376. https://doi.org/10.1016/j.compositesa.2003.09.016
[2] Aziz, S.H., Ansell, M.P. (2004). The effect of alkalization and fibre alignment on the mechanical and thermal properties of kenaf and hemp bast fibre composites: Part 1 – polyester resin matrix. Composites Science and Technology, 64(9): 1219-1230. https://doi.org/10.1016/j.compscitech.2003.10.001
[3] Islam, M.S., Kim, L.P., Nic, J.F. (2010). Influence of alkali fiber treatment and fiber processing on the mechanical properties of hemp/epoxy composites. Journal of Applied Polymer Science, 119(6): 3696-3707. https://doi.org/10.1002/app.31335
[4] Bourmaud, A., Baley, C. (2010). Effects of thermo mechanical processing on the mechanical properties of biocomposite fl ax fi bers evaluated by nanoindentation. Polymer Degradation and Stability, 95(9): 1488-1494. https://doi.org/10.1016/j.polymdegradstab.2010.06.022
[5] Liu, M., Fernando, D., Daniel, G., Madsen, B., Meyer, A., Ale, M.T., Thygesen, A. (2015a). Effect of harvest time and field retting duration on the chemical composition, morphology and mechanical properties of hemp fibers. Industrial Crops and Products, 69: 29-39. https://doi.org/10.1016/j.indcrop.2015.02.010
[6] Henriksson, G., Akin, D.E., Hanlin, R.T., Rodriguez, C., Archibald, D.D., Rigsby, L.L., Eriksson, K.L. (1997). Identification and retting efficiencies of fungi isolated from dew-retted flax in the United States and Europe. Applied and Environmental Microbiology, 63(10): 3950-3956. https://doi.org/10.1089/oli.1.1997.7.523
[7] Cabrol, L., Malhautier, L., Poly, F., Lepeuple, A.S., Fanlo, J.L. (2010). Assessing the bias linked to DNA recovery from biofiltration woodchips for microbial community investigation by fingerprinting. Applied Microbiology and Biotechnology, 85(3): 779-790. https://doi.org/10.1007/s00253-009-2253-8
[8] Mazian, B., Bergeret, A., Benezet, J.C., Malhautier, L. (2018). Influence of field retting duration on the biochemical, microstructural, thermal and mechanical properties of hemp fibres harvested at the beginning of flowering. Industrial Crops and Products, 116: 170-181. https://doi.org/10.1016/j.indcrop.2018.02.062
[9] Segal, L., Creely, J.J., Conrad, M., Martin, A.E. (1958). An empirical method for estimating the degree of crystallinity of native cellulose using the X-Ray diffractometer. Textile Reserach Journal, 786-794. https://doi.org/10.1177/004051755902901003
[10] Ribeiro, A., Pochart, P., Day, A., Mennuni, S., Bono, P., Baret, J., Spadoni, J., Mangin, I. (2015). Microbial diversity observed during hemp retting. Applied Microbiology and Biotechnology, 99(10): 4471-4484. https://doi.org/10.1007/s00253-014-6356-5
[11] Akin, D.E., Epps, H.H., Archibald, D.D., Sharma, H.S.S. (2000). Color measurement of flax retted by various means. Textile Reserach Journal, 70: 852-858. https://doi.org/10.1177/004051750007001002
[12] Liu, M., Ale, M.T., Kołaczkowski, B., Fernando, D., Daniel, G., Meyer, A.S., Thygesen, A. (2017). Comparison of traditional field retting and Phlebia radiata Cel 26 retting of hemp fibres for fibre-reinforced composites. AMB Express, 7: 58. https://doi.org/10.1186/s13568-017-0355-8
[13] Schäfer, T., Honermeier, B. (2006). Effect of sowing date and plant density on the cell morphology of hemp (Cannabis sativa L.). Industrial Crops and Products, 23(1): 88-98. https://doi.org/10.1016/j.indcrop.2005.04.003
[14] Nykter, M., Kyma, H., Belinda, A., Lilholt, H., Thygesen, A. (2008). Effects of thermal and enzymatic treatments and harvesting time on the microbial quality and chemical composition of fibre hemp (Cannabis sativa L.). Biomass and Bioenergy, 32(5): 392-399. https://doi.org/10.1016/j.biombioe.2007.10.015
[15] Meijer, W.J.M., Vertregt, N., Rutgers, B., van de Waart, M. (1995). The pectin content as a measure of the retting and rettability of flax. Industrial Crops and Products, 4(4):273-284. https://doi.org/10.1016/0926-6690(95)00041-0
[16] Musialak, M., Wróbel-Kwiatkowska, M., Kulma, A., Starzycka, E., Szopa, J. (2008). Improving retting of fibre through genetic modification of flax to express pectinases. Transgenic Research, 17(1): 133-147. https://doi.org/10.1007/s11248-007-9080-4
[17] Placet, V., Day, A., Beaugrand, J. (2017). The influence of unintended field retting on the physicochemical and mechanical properties of industrial hemp bast fibres. Journal of Materials Science, 52(10): 5759-5777. https://doi.org/10.1007/s10853-017-0811-5
[18] Kabir, M.M., Wang, H., Lau, K.T., Cardona, F. (2013). Effects of chemical treatments on hemp fibre structure. Applied Surface Science, 276: 13-23. https://doi.org/10.1016/j.apsusc.2013.02